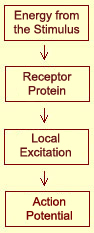
The steps that lead to the activation of a nociceptor
are the same as those for specialized tactile receptors. The energy of the pain
stimulus, which may be mechanical, thermal, or chemical, alters the conformation
of certain
proteins in the cell membranes of the A
delta or C fibres . This change in conformation modifies the permeability
of the membrane so as to induce a local excitation proportional to the energy
of the stimulus. Once this excitation reaches a
certain threshold, a nerve impulse, or action
potential, is transmitted.
And because the amplitude of
an action potential is always the same, the variations in the intensity of the
nociceptive stimulus will then be translated into the frequency of the train of
nerve impulses, which is the neurons’ preferred method of communicating
with one another.
At
each of the synapses
along this pain pathway, several neurotransmitters are involved in carrying the
nociceptive message. Those identified to date fall into two major groups: “classical”
neurotransmitters and neuropeptides.
Examples of classical neurotransmitters include glutamate,
aspartate, and serotonin. At least 20 neuropeptides involved in transmitting pain
impulses have been identified, including substance P, vasoactive intestinal peptide,
calcitonin gene-related peptide, somatostatin, cholecystokinin, and ACTH, not
to mention the enkephalins,
a large family of peptides that exert an inhibitory effect on the descending
control pathways.
A single nociceptive fibre can
contain a variety of different peptides and classical neurotransmitters, and their
respective roles remain largely undetermined. It is also hard to establish any
correlations between the kinds of peptides that the various nociceptive pathways
contain and their electrophysiological properties.
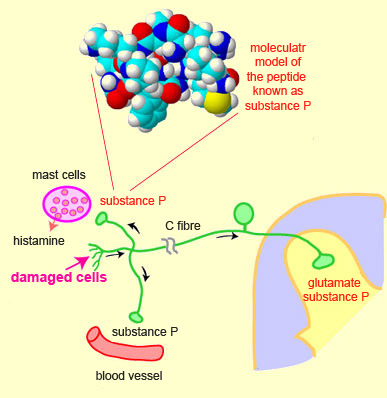
It is known, however, that glutamate
and substance P (a peptide containing 11 amino acids and belonging to the
tachykinin family) seem to be among the substances most involved in the transmission
of pain. For example, substance P binds to specific
receptors, called NK1 receptors, that are located on the nociceptive neurons
of the dorsal horn of the spinal cord.
Substance P also occurs in the brain, where it
is associated with regulation of mood disorders, anxiety, reinforcement,
neurogenesis, neurotoxicity, respiratory rate, nausea, and, of course, pain.
Also, through a phenomenon known as the axon reflex, substance P can be released
in the peripheral nervous system at the site of a tissue injury, where it causes
strong vasodilation that can induce the release
of various substances, such as bradykinin, histamine, and serotonin.
In
general, substance P has been associated with relatively slow excitatory connections,
and hence with the persistent, chronic pain sensations transmitted by C
fibres, whereas glutamate is involved in the rapid neurotransmission of acute
pain associated with A
delta fibres. The receptors for substance P and glutamate can be distributed
in different populations of neurons that preserve their own specific characteristics.
But the two types of receptors can also coexist on the same neurons, as has been
observed in several different parts of the central nervous system. Many pharmaceutical
companies have tried to develop substance P antagonists in hope of using them
as powerful analgesics, but the results have been very disappointing.
In
the peripheral nervous system, other
substances also contribute to the transmission of pain and make the nociceptors
more sensitive. Some of these substances, such as hydrogen and potassium ions,
arise from the tissue injury itself. Others, such as leucotrienes and prostaglandins,
are associated with the process of inflammation and act by sensitizing the nociceptors
to the substances generated by the injury. Still other substances, such as substance
P, are released by the nociceptors themselves and activate them directly.
In
parallel with the process of descending
control of pain and the endorphins
associated with it, which let you tolerate painful bodily effort and focus on
something other than pain, these processes of sensitization and inflammation tend
to make you immobilize the injured part of your body so as to facilitate the effects
of this “inflammatory soup” of molecules, along with the healing process.
The increased pain that even the simplest tactile stimuli produce around the sensitized
site of the injury provide a good incentive for you to take good care of the injured
part of your body.
The central sensitization that
occurs in the spinal cord can amplify the pain response to a normal stimulus even
further. This sensitization occurs through a different
set of cellular mechanisms.
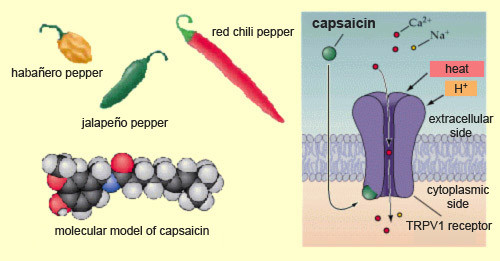
Nearly a third of the world’s people consume
hot peppers such as jalapeños every day. The “heat” in these
peppers comes mainly from capsaicin, a molecule that causes a burning sensation
by binding to special
receptors called TRPV1 receptors, located on the nociceptors. TRPV1
receptors can also be activated by heat or by an endogenous compound, anandamide,
which also activates the brain’s receptors
for cannabinoids. Heat and and anandamide are therefore probably
the natural activators of these TRPV1 receptors which, by chance, can also be
activated by an exogenous molecule from a plant, such as capsaicin.
Adapted from: Neuroscience, by Dale Purves
et al.
TRPV1 is a member of the vanilloid family of receptors.
Vanilloids are channel
receptors that, when stimulated, allow calcium and sodium ions
to enter the neuron. As a result, the neuron becomes depolarized, and if the depolarization
passes a certain threshold, it triggers action
potentials. The nociceptor then releases substance P, which excites
the next neuron in the ascending nociceptive pathway, and so on up to the brain,
at which point you realize just how spicy that food you’re eating actually
is!
And yet, a few seconds later, you often find that it’s not really
so spicy as you thought at first. The reason is that prolonged contact between
capsaicin and its receptors desensitizes them. Ironically, capsaicin can therefore
also produce analgesia, caused in part by the depletion of substance P. That is
why capsaicin is one of the main ingredients in certain analgesic and anti-inflammatory
creams that are used to relieve not only simple muscle and joint pain, but also
other forms of pain that are harder to treat, such as arthritis and neuropathic
pain. These creams often contain another ingredient such as lidocaine
to reduce the burning sensation that the capsaicin initially causes.
Capsaicin
receptors are found in all mammals, but not in birds, which has enabled manufacturers
to produce squirrel-proof seed for bird feeders! Research has also shown that
mice in which the gene for the capsaicin receptor has been deactivated can drink
a capsaicin solution as if it were ordinary water.
Phylogenetic studies have shown the importance
of the endorphins that are present in all vertebrates. Some scientists even believe
that these endorphins may have helped vertebrates free themselves from the automatic
protective reactions otherwise triggered by nociceptive stimuli, and thus helped
to encourage behaviours that are more adaptive in many situations.
For
example, if a wounded prey animal stopped to lick its wounds instead of continuing
to run from its predator despite the pain, its chances of survival would be limited.
But instead, the endorphins that the animal secretes in response to its fear,
stress, and the physical exertion of running make this pain bearable enough for
the animal to continue to flee.
Thus the endorphins let the animal give
priority to surviving, and take care of recovering and healing later on.
Subsequent studies, however, have made the picture
more complex. It now seems that the placebo effect has one component that is attributable
to endorphins and another that is not. The former is more associated with expectations
and the latter with conditioning.
Once endorphins have bound to their receptors
and produced their effects, they are quickly deactivated. The main mechanism by
which this deactivation takes place is that of a family of enzymes called peptidases,
which break the endorphins down by severing the bonds between the various amino
acids of which these opioid peptides are composed.
In 2003, researchers
isolated a substance called sialorphin that is secreted in rats. Sialorphin
binds to the enzymes that would otherwise break down enkephalins and thereby prevents
these enzymes from doing so. The enkephalins can then remain active longer, resulting
in a powerful pain-suppressing effect. When researchers injected rats with sialorphin,
they were able to walk freely over a bed of nails.
Under natural conditions,
sialorphin is released into the rat’s bloodstream in response to stress.
For example, when male rats are subjected to conditions of competition
and aggression among themselves, sialorphin reduces the pain that they
feel from the resulting injuries.
These interesting properties of rat sialorphin
led researchers to start looking for its functional analogue in humans. And within
a few years, an equivalent molecule, opiorphin, was discovered. Scientists
are now working to determine in what situations opiorphin is secreted in humans
and how it contributes to the analgesic effect of endorphins.
A better
knowledge of natural peptidase blockers such as sialorphin could also help researchers
to design new medications that could reduce pain by preventing the breakdown of
endogenous opioids.
Opium was most likely known as far back
as the time of the Sumerians (about 3 000 B.C.E.), to judge from written records
on their cuneiform tablets. Certain Egyptian documents from the reign of Ramses II
(1 300 B.C.E.) explicitly praise this plant’s abilities to induce sleep
and ease pain.
Opium
smokers in France, cover of Le Petit Journal, July 5, 1903
But
it was not until the 18th century that opium became a subject of scientific interest.
A first active ingredient of opium was described as a plant alkaloid by F. W.
Serturner in works published in 1805-1806 and 1817. Because of this substance’s
ability to induce sleep in human beings, Serturner named it morphium (in English,
morphine), after Morpheus, the god of dreams
in ancient Greece.
The
complex molecular structure of morphine was first described many years later,
in 1925, by British chemist Robert Robinson. Starting in 1952, scientists were
able to chemically synthesize morphine and its derivatives, eventually also producing
compounds whose structure was similar to morphine but whose effects differed somewhat
(for example, dextromethorphan, an analgesic similar to codeine). After that,
opiates very soon became widely used in medicine. For example, Henri
Laborit developed an injectable “cocktail” of opiates and tranquilizers
that was used to facilitate the transfer of wounded soldiers to operating rooms
during the French war in Indochina.
Chemically,
the body’s endogenous opiates are peptides—small
proteins that consist of short chains of amino acids and that are synthesized
right inside the nerve cells by
means of the cells’ messenger RNA and ribosomes, just like any other
proteins. (More specifically, all of these peptides are produced by the cleavage
of longer, “precursor” proteins.) These peptides are then carried
down the axons to the nerve endings, where they are released.
The
general term for these endogenous peptides is “endorphins”, a reference
to the similarity between their effects and those of morphine. At least 20 different
endorphins are known to be present in the human brain. The following paragraphs
describe the main categories of endorphins and the precursor proteins from which
they are derived.
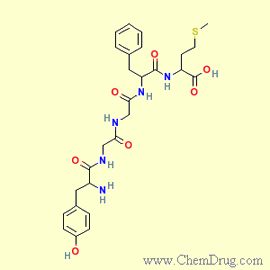
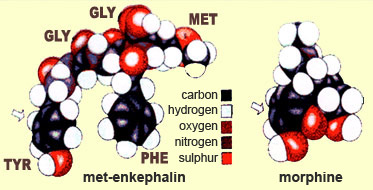
Enkephalins—more
specifically, met-enkephalin and leu-enkephalin—are
the first two endorphins to have been identified. Both of these peptides consist
of a chain of five amino acids, the first four of which are identical. The two
differ only in the last amino acid in the chain: methionine, in the case of met-enkephalin,
and leucine, in the case of leu-enkephalin.
Enkephalins
are produced by the cleavage of a precursor protein called proenkephalin. Every
proenkephalin molecule contains at least seven active peptide molecules, including
four met-enkephalin molecules and one leu-enkephalin molecule. Once the proenkephalin
molecules have been cleaved by what are known as maturation enzymes, these enkephalin
molecules are released.
A
comparison of the structures of an endogenous opiate and morphine shows that the
two molecules have one area that is similar. This explains why they share an affinity
for the opioid receptors in the brain.
Enkephalins
are secreted in all the structures of the central and peripheral nervous systems,
close to the mu
and/or delta opioid receptors that are their natural receptors. Shortly after
they have done their job of modulating pain, these natural opioids are deactivated
when they are cleaved by a family of enzymes known as metallopeptidases.
Dynorphins
are a class of endogenous opioids that play a powerful role in modulating pain.
(Their power is reflected in their name, which, like the word “dynamic”,
comes from the Greek dynamis.)
Dynorphins
are derived from the precursor protein prodynorphin. When prodynorphin is cleaved
by the enzyme proprotein convertase 2 (PC2), several active opioid peptides are
released, including dynorphin A, dynorphin B, and alpha and beta neoendorphin.
These four peptides contain the exact same sequence of amino acids as leu-enkephalin,
but with additional amino acid molecules as well (12, 8, 5, and 4, respectively).
Dynorphins are distributed broadly in the
central nervous system, but are found in especially high concentrations
in the hypothalamus, the brainstem, and the spinal cord. Their physiological
effects differ with the site where they are produced, and they bind
mainly to kappa
opioid receptors (though they also have a strong affinity for
mu and delta receptors).
The term endorphin refers not only to endogenous
opioid peptides in general, but also to a specific group of such
peptides in particular. These endorphins are distinguished by a
Greek letter at the start of their name. The most important of these
is beta-endorphin, which, in addition to reducing pain substantially
(its analgesic power is several times greater than that of morphine),
is also the opioid peptide that produces the greatest sensation
of euphoria. Beta-endorphin is produced in large amounts during
sustained
physical exercise and produces this sensation by binding to
mu
opioid receptors.
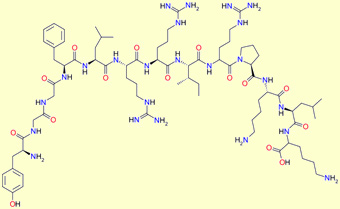
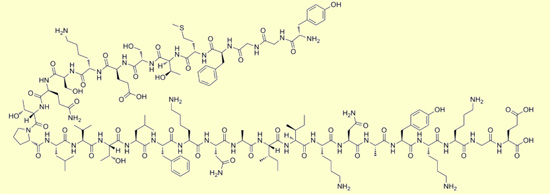
Beta-endorphin
Beta-endorphin: Tyr Gly Gly Phe Met
Thr Ser Glu Lys Ser Gln Thr Pro Leu Val Thr Leu Phe Lys Asn
Ala Ile Ile Lys Asn Ala Tyr Lys Lys Gly Glu
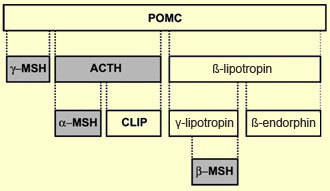
The precursor of
beta-endorphin, pro-opiomelanocortin (POMC), is unusual in that
its cleavage yields not only beta-endorphin and other opioid peptides,
but also several other peptide hormones. Depending on the type of
tissue where the gene for POMC is expressed, it will be cleaved
differently and produce different peptides.
Thus, in the anterior pituitary gland,
POMC yields not only beta-endorphin and beta-lipotropin (an opioid peptide associated
with fat metabolism) but also adrenocorticotropic
hormone (ACTH), a hormone secreted in response to stress. But when the gene
for POMC is expressed in the melanocytes of the skin, POMC instead yields the
hormone melanotropin, which triggers the synthesis of the pigment melanin, which
causes the skin to darken in response to the sun’s rays.
Summary
of the possible derivatives of POMC
POMC,
which is also found in the hypothalamus, is a chain of 241 amino acids and can
be split at various points by enzymes called prohormone convertases. One of its
derivatives, beta-lipotropin, has 90 amino acids and was first isolated in 1964
by biochemist C. H. Li. Initially, Li could not determine what its function might
be, but after seven years he discovered that beta-lipotropin was, among other
things, a precursor of beta-endorphin. Beta-endorphin comprises 31 amino acids,
making it the longest member of its family, which also includes alpha-, gamma-,
and sigma- endorphin.
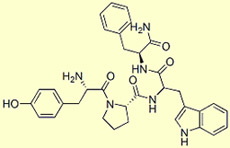
Endomorphin
1
Endomorphin 1: Tyr-Pro-Trp-Phe-NH2
Endomorphin
2: Tyr-Pro-Phe-Phe-NH2
The neurons
of the hypothalamus also contain another group of opioid peptides discovered in
the late 1990s: endomorphin1 and endomorphin
2, two small peptides that have four amino acids and the greatest known
affinity with the mu
opioid receptors.
The name endomorphin (not
endorphin, as in beta-endorphin) is a reminder that these proteins are produced
naturally within our own bodies and bind to the same receptors that enable exogenous
substances such as morphine to produce their effects.
The
anatomical distribution of endorphins in the body suggests that they play a role
in pain control, responses to stress,
wakefulness,
and the reward
circuit.
Also
in the late 1990s, another opioid peptide was discovered: orphanin FQ (or
nociceptin), which has 17 amino acids, is derived from the precursor prepronociceptin,
and binds to only a very limited extent to classic opioid receptors. Instead it
prefers an atypical class of opioid receptors called orphan
receptors (whence its name).
The mechanism by
which orphanin is involved in the perception of pain is complex, because it can
act as an analgesic at some times and an anti-analgesic at others (by blocking
the effects of other opioid peptides).
One last opioid peptide that should be mentioned
here is nocistatin, which also is derived from the precursor prepronociceptin
and is apparently involved not only in the transmission of pain, but also in memory
and learning.