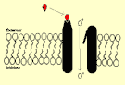
ionotropic receptors are
ion channels to which neurotransmitters bind directly in
order to open them.
Click to enlarge
In contrast, metabotropic
receptors are separate from the ion channels whose
operation they regulate. They make the linkage by means
of a membrane protein from the G-protein family.
Click to enlarge
SYNAPSES
Thanks to research by hundreds of laboratories
throughout the world, the main entities involved in synaptic transmission
have now been identified. They include over sixty neurotransmitters
and hundreds of subtypes of receptors. Since a single neuron may
release several different neurotransmitters at once, the soup of
molecules and ions in the synaptic gap can be decoded only by means
of very specific affinities between neurotransmitters and their
receptors.
A combination of neurotransmitters that can
act on various subtypes of receptors can thus have varying effects,
depending on what particular receptors they are acting upon.
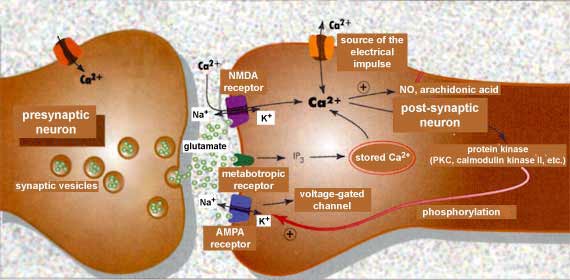
This diagram shows a synapse that is capable
of long-term
potentiation, a synaptic facilitation mechanism that is the
basis for memory. The neurotransmitter involved here is glutamate.
The enlarged diagram shows just three of the approximately twenty
known subtypes of glutamate receptors.
When glutamate binds to these receptors,
it not only causes the ion channels to open but also triggers several cascades
of chemical reactions, also represented in this diagram. Many
of these reactions involve “second messengers:” molecules
that relay signals from neurotransmitters within the postsynaptic
neuron and that can in turn cause other ion channels to open or
close. The effects of second messengers can extend all the way
to the neuron’s nucleus, thus influencing its synthesis of
new proteins, receptors, or channels, for example.
Ion channels
too are large proteins embedded in the neuronal membrane.
It is the selective opening of all these various channels
that, by changing the membrane’s electrical potential,
produces the action
potential.
It is also these channels that let
calcium ions enter the presynaptic neuron when the action
potential reaches the axon’s terminal button–a
crucial step that leads to the fusion of the synaptic vesicles
with the membrane and their expulsion of neurotransmitters
into the synaptic gap.
Lastly, this overview of the entities
involved in neurotransmission would
not be complete without a mention of the other transmembrane
proteins that reabsorb neurotransmitters into the presynaptic
neuron or that actively pump ions through the membrane against
their natural gradient.
Many peptides act more as neuromodulators than
as neurotransmitters. Neuromodulators are substances that
do not propagate nerve impulses directly, but instead affect
the synthesis, breakdown, or reabsorption (reuptake) of neurotransmitters.
Neuromodulators can also exert regulatory effects on many
extra-synaptic receptors, rather than on synaptic sites exclusively.
NEUROTRANSMITTERS
To be considered a neurotransmitter, a molecule must meet several
criteria.
1) It must be produced inside a
neuron, found in the neuron’s
terminal button, and released into the synaptic gap upon the
arrival of an action potential. 2) It must produce an effect
on the postsynaptic neuron. 3) After it has transmitted its signal
to this neuron, it must be deactivated rapidly. 4) It must have
the same effect on the postsynaptic neuron when applied experimentally
as it does when secreted by a presynaptic neuron.
Over 60 different molecules are currently known to meet these
criteria.
Among the small molecules constituting
the “classical” neurotransmitters,
the best known are:
acetylcholine
serotonin
catecholamines, including epinephrine, norepinephrine,
and dopamine
excitatory amino acids such as aspartate and glutamate
(half of the synapses in the central nervous system
are glutamatergic)
inhibitory amino acids such as glycine and gamma-aminobutyric
acid (GABA; one-quarter to one-third of the synapses
in the central nervous system are GABAergic)
histamine
adenosine
adenosine triphosphate (ATP)
Peptides form
another large family of neurotransmitters, with over
50 known members. Here is a very partial list:
Certain soluble
gases also act as neurotransmitters. The most
important member of this category is nitrogen monoxide
(NO).
These neurotransmitters act by their own distinctive
mechanism: they exit the transmitting neuron’s
cell membrane by simple diffusion and penetrate the
receiving neuron’s membrane in the same way.