|
|
| Funding for this site is provided by readers like you. | |
|
|
|
|
|
|||||
|
|
|||||||
|
|
|
|
|
|
|
|
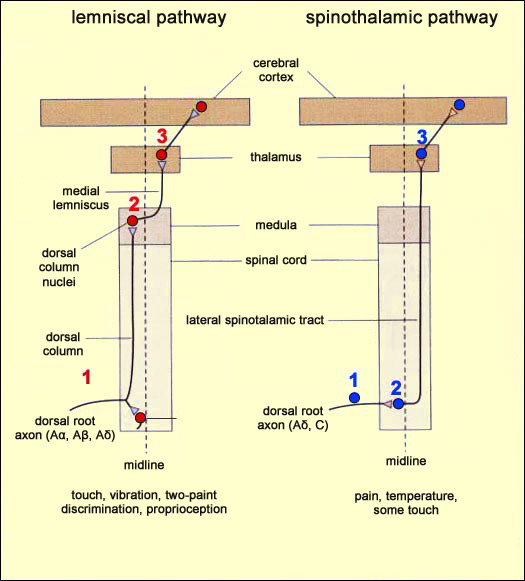
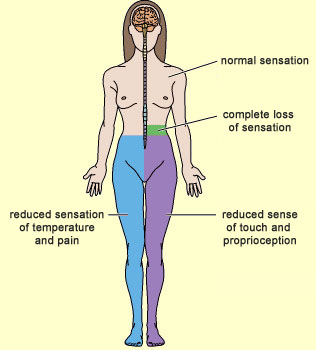
What pathways do pain signals follow to perform their protective function? How do these messages reach your brain to tell you which part of your body is hurt? As you might expect for a function as essential as the sensing of pain, the pathways for these signals are numerous, complex, and mutually redundant. But before we can trace these pain pathways (also known as nociceptive pathways), we must distinguish them from the sensory pathways for non-painful temperature, touch, and proprioception. These various sensory signals take two different paths to reach the brain, both of which start in a given part of the body and end in the brain’s somatosensory cortex. Each of these paths consists of a chain of three neurons that pass the nerve impulses from one to the next. Where these two paths differ is in the location where they cross the midline in the spinal cord. Remember that in the human body, the nerves responsible for sensory inputs, as well as those responsible for motor control, are crossed. In other words, the neural pathways from the left side of the body terminate in the right hemisphere of the brain, and vice versa. Hence, at some point in the body, these pathways must cross the body’s midline (in scientific terminology, they must “decussate”). Now let’s follow the path that any incoming sensory impulse—whether for touch, pain, heat, or proprioception—follows from the spinal cord to the brain. Regardless of the sensory modality, the three neurons in question form a chain running from one side of the spinal cord to the other, and the cell body of the first neuron in this chain is always located in a spinal (dorsal root) ganglion. This neuron is said to be T-shaped, because its axon emerges as a short extension from its cell body and then soon divides into two branches going in opposite directions: one goes to the part of the body that is innervated by this spinal nerve, while the other immediately enters the dorsal root of the spinal cord (an essentially sensory part of the spinal cord, as opposed to the ventral root, which is a motor area). It is from this point on that the two pathways differ.
Adapted from Neuroscience: Exploring the Brain, M.F. Bear, B.W. Connors, and M.A. Paradiso, 2007
In both of these pathways, the third neuron sends its axon to the somatosensory cortex, the part of the brain that determines exactly where the original stimulus occurred in the body.
|
|
||||||||||||||||||||||||||||||||||||

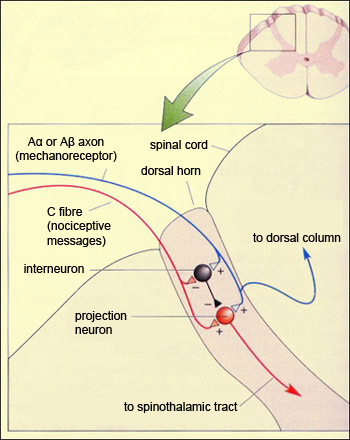
The perception of pain results not simply from the activation of the ascending nociceptive pathways, but from an actual dialogue between these pathways and the various descending pathways that control this pain. The control mechanism involved is often described as a system of filters or a set of gates whose closing is controlled by the cortex, the midbrain, and the medulla. But the incoming nociceptive impulse encounters its very first gate as soon as it enters the dorsal root of the spinal cord. This first relay point in the ascending pathway is thus not just an area through which the nociceptive impulse passes, but rather the first place where it is filtered and integrated with other information. This first level of integration is referred to as segmental controls of non-pain peripheral origin. The word “segmental” refers to the fact that this process occurs in each of the segments of the spinal cord corresponding to each vertebra. This segmental control results from the interaction between the nociceptive sensory fibres (A delta and C) and the non-nociceptive ones (A alpha and A beta). This interaction was modelled in an article, first published in 1962 and then amplified in 1965, that many regard as the most important one ever written on the subject of pain. In this article the authors, Canadian Ronald Melzack and Englishman Patrick Wall, proposed the first model for the endogenous control of pain: the now-famous gate control theory of pain.
This theory posits a special form of connectivity involving not only the sensory input fibres for pain and for light touch, as mentioned above, but also a set of inhibitory interneurons
that are the key element in the authors’ explanation.
It is this particular circuit that forms the virtual gate whose opening and closing will modulate the passage of pain. Under normal conditions, the inhibitory interneurons spontaneously produce action potentials at their own specific frequency. But when the nociceptive fibres are activated by a pain stimulus, in addition to stimulating the non-specific neuron that projects to the spinothalamic pathway (also known as the “projection neuron”), they also inhibit the spontaneous inhibitory activity of the interneurons, thus depolarizing the projection neuron and increasing the likelihood that it will trigger action potentials. Another aspect of this circuit’s operation is illustrated by what happens when you hurt yourself and start to rub the injured part of your body vigorously. This instinctive reaction reduces the sensation of pain by “closing the gate”. The animation below shows how: rubbing your skin activates the sensory fibres for touch, which in turn excite the projection neuron. But these fibres also make numerous excitatory connections to the inhibitory interneurons. As a result, if you keep rubbing your skin, these interneurons will produce a strong hyperpolarization of the projection neuron, thus greatly reducing the probability that it will emit nerve impulses.
Thus we see how it is the relative frequencies of the action potentials in the nociceptive and non-nociceptive fibres that determine how open the “gate” in the spinal cord will be and hence how much pain information will pass through. In addition, there are projections of central origin that can also activate these inhibitory interneurons in the spinal cord and further close the gate at the segmental level. Data gathered since 1965 have led to some changes in Melzack and Wall’s original model, but the idea that the perception of pain is modulated from the moment that the pain messages enter the spinal cord remains fundamental to the clinical treatment of pain. For example, it is the origin of clinical applications such as transcutaneous electrical nerve stimulation (TENS), which produces local analgesia by stimulating the non-nociceptive fibres in the skin.
|
| |
|
|
|
|
|
|
|
|