The
100 trillion cells in the adult human body are divided
into 300 different types of cells, each of which expresses
only a subset of the genome specific to its type. Amazingly,
only 1.1% of the DNA in our genes represents instructions
for manufacturing proteins. So how does the human body
manufacture the tens of thousands of proteins that it contains,
if it has only around 30 000 genes with which to do it?
Obviously, a “one gene, one protein” model
will not provide the answer.
The answer lies in what are known as post-transcriptional processes.
For example, the “gene for insulin”, the hormone
needed to metabolize sugar, actually encodes for the manufacturing
of a very large protein that will subsequently be broken down
into smaller pieces by enzymes. And it is the combination (or “splicing”)
of two of these pieces that yields insulin. In all, this single
gene is used to produce four different proteins and three different
hormones.
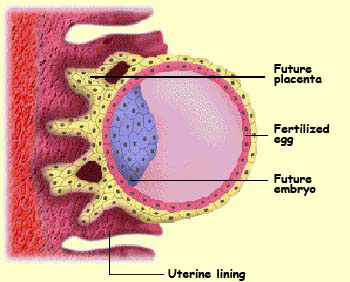
When the embryo becomes
deeply embedded in the uterine wall, openings form in the
layer of external cells and fill with the mother’s
blood. This is the embryo’s first contact with the
source that will nourish it throughout its development. The placenta is
thus composed both of trophoblasts from the embryo and of
endometrial cells from the mother’s uterus.
Following implantation, some major hormonal adjustments occur.
The hormone human chorionic gonadotropin (hCG)
is secreted by the embryo’s trophoblasts and stimulates
the production of progesterone and estrogen by
the ovaries (hCG is the hormone that is detected in pregnancy
tests). From the 10th to the 14th day, the amniotic
cavity (which protects the fetus from external shocks)
and the placenta (where exchanges take place between the fetus
and the mother) begin to form. At this stage, the mother has
probably just noticed that she has missed her period.
Implantation normally
takes place around the 10th day, in the upper third of the
uterus, along the median line. When the embryo becomes implanted
somewhere other than the uterus, it is usually in one of
the fallopian tubes. This abnormal condition is called an ectopic
pregnancy.
FROM FERTILIZATION
TO EMBRYO
Sexual reproduction in human beings is made
possible by the fusion of two reproductive cells, or “gametes”—the ovum
from the mother and the sperm cell from the father. This fusion
results in the formation of a zygote with 46 chromosomes:
half from the mother and half from the father. The zygote thus
acquires a unique genetic identity, so that it is far more accurate
to speak of the sex act as one of procreation (the creation of
a new being) rather than one of reproduction, which would imply
simply producing the same thing over again.
One day after fertilization,
the zygote begins a series of mitotic
divisions in which the considerable volume of cytoplasm from
the ovum is divided among the multiple daughter cells. Hence, during
these first cell divisions, the zygote, now called an embryo, does
not grow in size but does move successively through the stages
where it is composed of 2, 4, 8, and then 16 cells, called blastomeres.
This first phase of development, in which the embryo undergoes
a series of rapid divisions in close succession, is called segmentation.
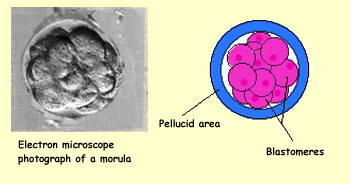
In humans, the embryo reaches the 16-cell
stage, also called the morula, on the third day
after fertilization. At this stage, the embryo is a compact ball
of cells that descends through the fallopian tube toward the uterus.
The cell divisions still have not caused the embryo to grow any
larger than the original ovum.
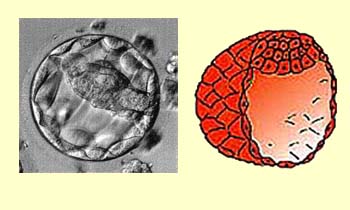
At the blastula stage, starting
at 128 cells, the ball of cells develops an inner cavity called
the blastocoele. This stage begins around the fourth day and continues
until the embryo has become implanted in the wall of the uterus,
around the sixth or seventh day. During this stage, the protective
envelope (or pellucid area) around the ball of
cells gradually decays, precisely so that the embryo can attach
itself to the uterine wall. It is also at this stage that the embryo
differentiates into two kinds of cells: outer cells called trophoblasts, which
will contribute to the placenta, and inner cells called embryoblasts,
which will form the embryo as such.
Implantation (or nidation)
in the uterine wall begins around the seventh day, when the trophoblasts,
freed from the pellucid area, secrete an enzyme that lets the embryo
burrow into the uterine wall. As the trophoblasts proliferate,
they form two distinct layers of cells: one that continues to surround
the embryo, and another whose membranes fuse, forming a multinucleate
mass called the syncytium, which is responsible
for penetrating the uterine wall.
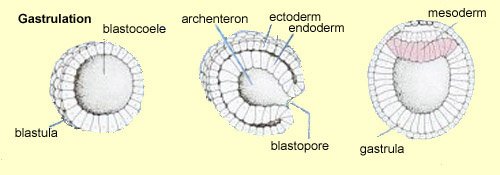
The third week of development begins with
a major cellular reorganization called gastrulation.
In gastrulation, a portion of the cells on the blastula’s
surface invaginates (penetrates inward), thus forming the endoderm,
while the cells remaining on the outside of this sphere form the
ectoderm. The cells inside the sphere then divide into two sheets
of cells forming two superimposed discs. The upper disc will become
the embryo, while the lower one will turn into a yolk sac that
provides nutrients to the embryo until a functional circulatory
system develops.
At the start of this phase, a narrow row
of cells traces a furrow along the embryonic disc and thus defines
the general axis around which the body’s entire bilateral
structure will develop. We can say that the embryo as such officially
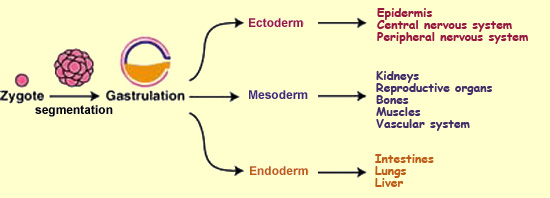
begins to exist starting at this stage, when a third layer of cells,
called the mesoderm, interposes itself between
the two that are already present, the ectoderm and the endoderm.
All of the cells in these layers have the same genetic material,
but some of them start to express certain genes rather than others
so as to develop particular organs. The cells in the innermost
layer, the endoderm, will produce such organs
as the intestines, the lungs, and the liver. The middle layer,
the mesoderm, will produce the kidneys, the reproductive
organs, the bones, the muscles, and the vascular system. And the
outer layer, the ectoderm, will be the source
of both the epidermis and the entire central and peripheral nervous
systems.
Source: adapted from BrainConnection.com
The co-ordinated movements that make
gastrulation possible are called morphogenetic movements,
and they involve the entire embryo. The cells adopt new positions
and consequently acquire new neighbours. Through gastrulation,
the cells are thus assembled into subgroups that act on one another
through induction
phenomena.
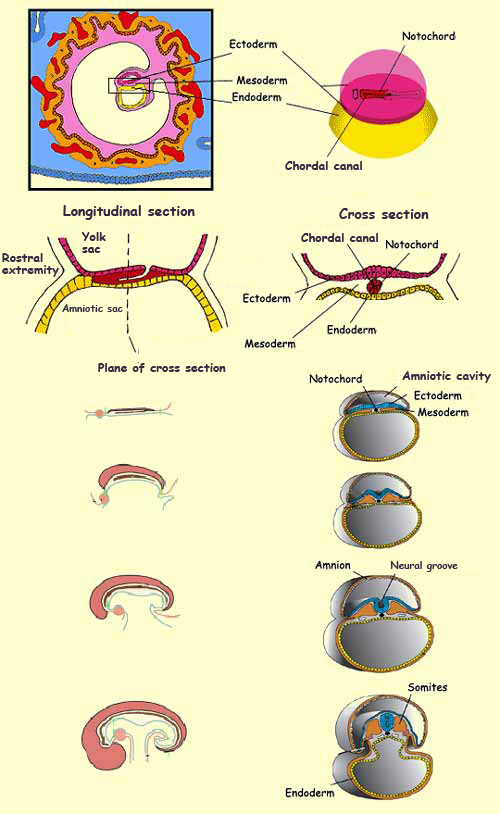
To sum up, gastrulation follows the rule
of threes: in the 3rd week, 3 cell layers form that
are the origin of 3 important structures: the primitive
streak, which defines the plane of bilateral symmetry
of the future embryo, and the dorsal cord (or notochord),
which will induce the formation of the neural plate in
the following stage, neurulation.
From six to eight weeks
after fertilization, the cerebral hemispheres begin to form.
Around the seventh week, nerves make connections with some
muscles that enable the embryo to make spontaneous movements.
At the end of the eighth week, all of the body’s essential
internal and external structures are present. The second and
third trimesters of pregnancy are devoted essentially to growth
of these structures that have already been put in place.
In the epithelium of
the neural tube, proliferation takes
place at specific locations called germinal zones. The germinal
zones from which most parts of the nervous system will develop
are located near the surface of one of the cavities that
will become the ventricular
system of the brain.
HOW THE NERVOUS
SYSTEM BEGINS
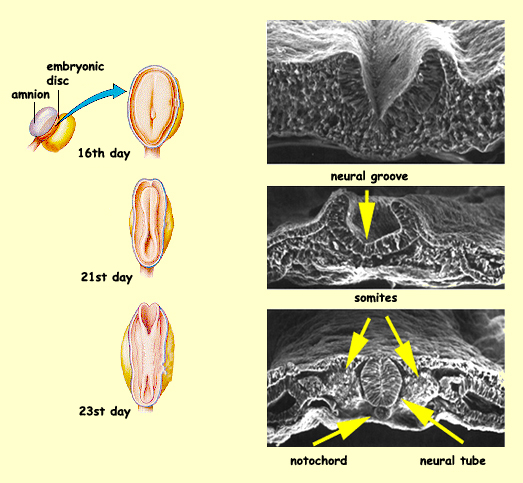
About three weeks after conception,
the human brain is nothing more than a single layer of flattened
cells located in the ectoderm and
known as the neural plate. Next, a furrow
forms that extends from the rostral portion to the caudal portion
of this plate. The sides of this neural furrow then
form the neural groove. The sides of this
groove then close over, starting from the middle of the groove
and moving outward rostrally and caudally, to form the neural
tube. Certain cells in the dorsal portion of the neural
tube will become the neural
crest, the structure that is the origin of the neurons
of the peripheral nervous system.
The part of the neural plate located just above the notochord
differentiates into the floor plate. The inductive
signals from this floor plate induce the development of the spinal
motor neurons and the motor neurons of the medulla and the
pons from the most ventral cells of the neural tube. The most
dorsal cells will give rise to the sensory neurons.
The process of formation of the neural
tube, which often begins before the mother even knows she is
pregnant, is called neurulation. It is from
this tube that the brain and the spinal cord will develop. At
this stage they will be the largest organs in the embryo, resulting
in its characteristic curved form. At the end of the third week,
the eyes and
the ears will also have begun to form.
After the segmentation and gastrulation phases
are completed, organogenesis begins. In this
phase, the groups of cells are laid down that will become the
various organs of the human body. Organogenesis starts with a
process called metamerization, in which the
mesoderm divides into a series of identical segments called metameres
along the embryo’s longitudinal axis. At this stage, the
mesoderm develops masses called somites on either
side of the neural tube. It is from these somites that the 33
vertebrae of the spinal column and the corresponding skeletal
muscles will develop.
Source: Dr. K. Tosney, University
of Michigan
At the start of the 4th week after fertilization,
the neural tube closes entirely, completing the first stage of
the development of the brain and the spinal cord. The next stage, histogenesis,
in which the stem
cells differentiate to form the various nerve tissues, can
now begin in earnest. At the same time, the
major subdivisions of the brain will form, and the cell populations
will be rearranged accordingly.
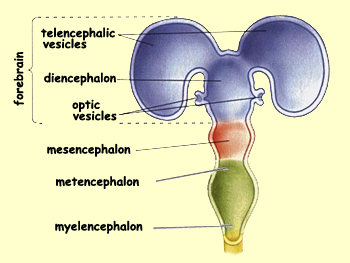
The telencephalon
is the most rostral of the secondary vesicles. Two buds
emerge from either side of its rostral portion to form
the two telencephalic vesicles. These two vesicles grow
rapidly to form the two cerebral hemispheres. First they
grow back over the diencephalon, then they grow down to
cover its sides.
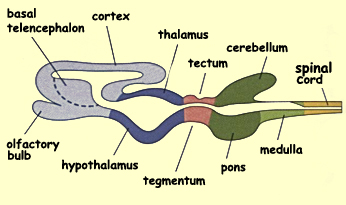
General diagram of a sagittal
section of the brain (applicable to all mammals)
Another pair of vesicles
will also sprout from the ventral surface of these cerebral
hemispheres to become the olfactory bulbs and
other structures that contribute to the sense of smell. Various
structures will then emerge from the walls of the telencephalon
while the white matter that connects these structures develops
as well. The neurons of the telencephalon wall proliferate
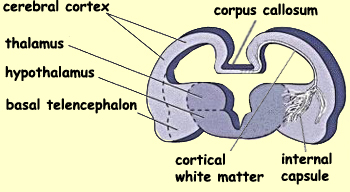
to form three distinct regions—the cerebral
cortex, the basal telencephalon, and
the olfactory bulb.
The axons of these
neurons will also gradually elongate to make connections
with the other parts of the nervous system. Some of these
axons will constitute the cortical white matter that
arises from and projects to neurons in the cortex. Others
will form the corpus callosum, the band
of nerve fibres that connects the two hemispheres of the
brain. Still others—those of the internal
capsule—will connect the cortical white
matter to the brain stem, generally by way of the thalamus.
For example, the axons
arising from the motor cortex will pass through the
internal capsule to connect to the motor neurons in the
spinal cord.
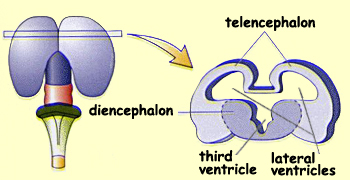
In the the remaining space between the telencephalon and
the diencephalon on either side, the two cerebral
ventricles (also known as the lateral ventricles or the
first and second ventricle) form, while the third ventricle
forms in the space
at the centre of the diencephalon.
The diencephalon also differentiates into distinct areas:
the thalamus and the hypothalamus.
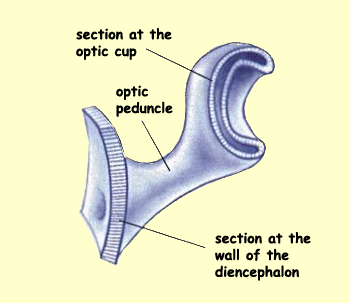
On either side of
the diencephalon, two secondary vesicles also develop—the optic
vesicles. The optic vesicles lengthen and fold
inward to form the optic peduncles and optic cups, which
will give rise to the retinas and
the optic
nerves. The retinas and the optic nerves are therefore
not part of the peripheral nervous system, but rather they
are integral parts of the brain!
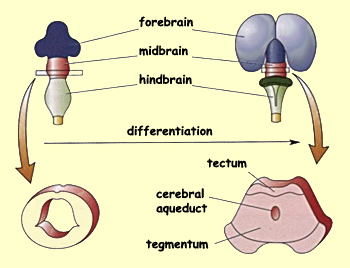
Compared with the prosencephalon (telencephalon and diencephalon),
the mesencephalon undergoes far less transformation. Its
dorsal surface forms the tectum,
while its floor forms the tegmentum. While
these structures are differentiating, the cavity that separates
them shrinks to a narrow channel called the cerebral aqueduct.
The rostral portion of this aqueduct opens into the third
ventricle of the diencephalon.
The mesencephalon serves as the passageway
for the bundles of fibres that connect the cortex to the spinal
cord—both those that arise from the sensory system and
those that descend to participate in movement
control.
The tectum differentiates into two structures. One, the superior
colliculus, receives information directly
from the eye and controls eye movements. The other, the inferior colliculus,
receives information from the ear and serves as an important
relay in the auditory pathways.
The tegmentum is one of the most colourful areas of the brain.
It contains the substantia nigra (“black
matter”) and the red
nucleus, two structures that are involved
in controlling voluntary movement. Other groups of cells in the
mesencephalon project
their axons diffusely into large areas of the brain and influence
a wide variety of functions, such as consciousness, mood, pleasure
and pain.
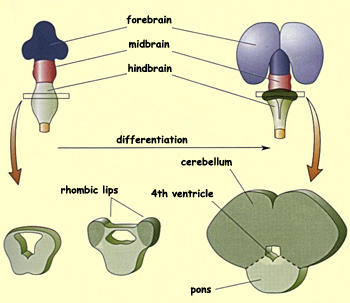
Caudal
to the mesencephalon lies the metencephalon, which is the
rostral portion of the hindbrain and differentiates into
two major structures: the cerebellum and
the pons. The cerebellum arises
from the thickening of the tissue covering the lateral
walls of the neural tube at this location. The two masses
thus formed ultimately fuse dorsally to form the cerebellum.
During this time, a swelling develops on the ventral side
of the metencephalon and forms the pons. This structure
is an important information pathway between the brain,
the cerebellum, and the spinal cord.
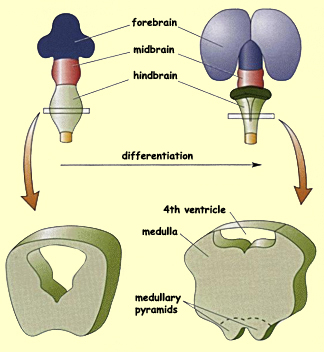
In the the myelencephalon (the caudal portion of the hindbrain)
the changes are less spectacular. The ventral and lateral
regions of this structure swell to form the medulla
oblongata. Along the ventral aspect of the medulla,
the two medullary
pyramids will also develop, formed by the passage of
the corticospinal bundles responsible for voluntary movement.
Lastly, the central canal, which persists while the medulla
is forming, becomes the fourth ventricle.
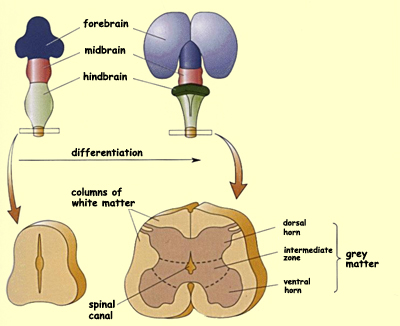
The entire portion of
the neural tube that lies caudal to the five secondary vesicles
becomes the spinal
cord through a fairly direct process of differentiation
consisting in the thickening of the tube walls. This thickening
gradually reduces the diameter of the neural tube until it
becomes the very narrow spinal canal .
As the cross-section shown here illustrates, the cell bodies
of the neurons in the spinal cord are concentrated in the grey
matter at the centre (the butterfly-shaped area),
while the white matter at the periphery is
composed of bundles of axons.
The grey matter of the spinal cord is in
turn divided into the dorsal horn, which receives
sensory inputs, and the ventral horn, whose
neurons innervate the skeletal muscles. Likewise, within the
white matter, there develop dorsal columns composed
of sensory axons that ascend to the brain and lateral
columns composed of corticospinal axons that descend
to transmit signals for controlling movement. Between the dorsal
and ventral horns, a large number of interneurons also develop
that are involved in various types of reflexes as well as in
establishing networks that perform initial processing of the
information received in the spinal cord.