Visual acuity is the eye's ability
to distinguish two points that are very close to each other. This ability depends
on many factors, but especially on the precision of the eye's refraction
and the ratio
of cones to rods at a given location on the retina.
THE EYE
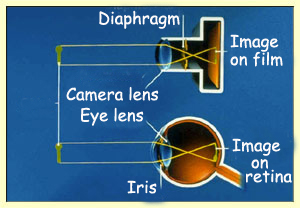
Functionally,
the eye can be compared with a camera, and the retina
with photographic film. The purpose of the camera is to focus an image that is
sharp and neither too dark nor too light onto the film. The photographer uses
the camera's focus ring to bring the image into focus and its diaphragm to ensure
that the amount of light entering the camera is just right for the sensitivity
of the film being used.
Your
eye does exactly the same thing, all day long, without your even being aware of
it! Your
cornea and lens provide the focus, while the iris adjusts to let the optimal
amount of light reach your retina. But your retina, with its many
layers of neurons, is far more complex and sensitive than any photographic
film. The two are similar, however, in that the image focused on both of them
is inverted.
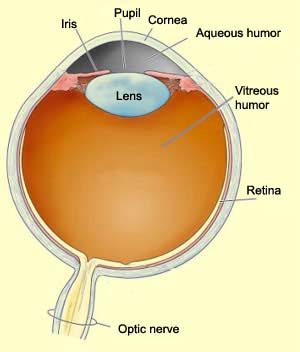
The main optical components of the
eye are as follows. First comes the cornea, the transparent,
slightly convex outer surface at the centre of the eye. The cornea does not have
any blood vessels, so its takes its nutrients from the fluid behind it, known
as the aqueous humour, as well as from the fluid in front of
it, the tears, which are spread across your cornea when you blink your eyelid.
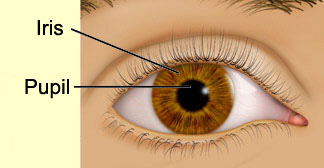
Next
comes the pupil, the opening that lets light enter the eye and
ultimately reach the retina. The pupil appears black because of the layer of black
pigmented cells that line the back of the eye and absorb the light.
The diameter of the pupil is controlled by the iris, a circular
muscle whose pigmentation gives the eye its colour and whose contraction lets
the eye adapt continuously to changing light conditions. On a dark night, your
pupils are big and black, because your irises open wide to let in as much as possible
of the little light available. This reaction is called the pupillary reflex.
You can observe it easily yourself, by watching your eyes in a mirror while you
turn a nearby light on and off.
After passing through the pupil, the
light goes on through the lens, which is suspended between the
aqueous humour and the vitreous humour, the fluid that fills
the inside of the eye.
The lens in turn focuses the light rays onto the
retina, lining the back of the eye. The retina converts the image formed by the
light rays into nerve impulses. The optic nerve, composed of the axons of the
retina's
ganglion cells, then transmits these impulses from the eye to the first
visual relay in the brain.
THE TARGETS OF THE OPTIC NERVE
The axons of the retina's
ganglion cells collect in a bundle at the optic
disc and emerge from the back of of the eye to form the optic nerve.
The optic nerve is the pathway that carries the nerve impulses from each eye to
the various structures in the brain that analyze these visual signals.
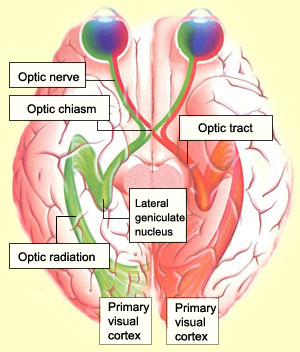
The optic nerves of the two eyes emerge
from their optics discs and intersect at the optic chiasm just
in front of the pituitary gland. In the optic chiasm, some of the axons from the
two retinas undergo decussation:
they switch sides to allow crossed processing of the visual signals.
The axons from the nasal side of each retina cross sides in the optic chiasm so
that the left
half of the field of vision is perceived by the right cerebral hemisphere,
and vice versa. But because the visual information that reaches the temporal side
of each retina comes from the opposite side of the visual field to begin with,
the axons from this side of the retina do not need to cross sides. Instead they
proceed straight ahead through the optic tract.
The vast majority of the nerve fibres in the optic
tract project to the lateral
geniculate nucleus (LGN) in the dorsal part of the thalamus. The LGN is the
main relay in the pathway to the primary visual cortex. The projection from the
LGN to the visual cortex is called the optic radiation. Because
damage at any point along the pathway from the retina to the cortex results in
some degree of blindness, this is clearly the pathway through which conscious
visual perception takes place in human beings.
People whose primary visual cortexes
have been damaged consider themselves to be blind and unable
to discern anything in their visual environment. But if you
ask these people to "take a chance" and point their
finger at a dot of light in space, they will point straight
at this target. And the data show that this result is not
random. This phenomenon is called blindsight.
Thus these
people are still processing some visual information, even though part of the neural
pathways in V1 have been destroyed. The mechanisms by which they do so may involve
little understood transfer pathways that bypass V1, as well as certain
subcortical visual nuclei. Some researchers also believe that the dorsal
visual pathway plays a role in this phenomenon.
THE VARIOUS VISUAL CORTEXES
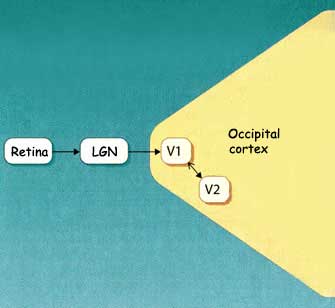
The image captured by
each eye is transmitted to the brain by
the optic nerve. This nerve terminates on
the cells of the lateral
geniculate nucleus, the first relay in the brain's visual pathways. The cells
of the lateral geniculate nucleus then project to their main target, the primary
visual cortex. It is in the primary visual cortex that the brain begins
to reconstitute the image from the receptive
fields of the cells of the retina.
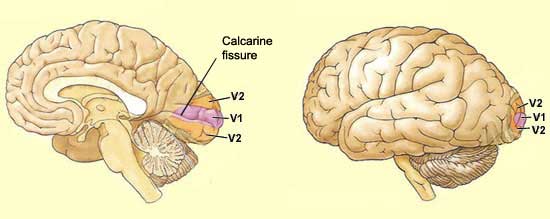
Also
known as the striate cortex, or simply V1,
the primary visual cortex is located in the most posterior portion of the brain's
occipital
lobe . In fact, a large part of the primary visual cortex cannot be seen from
the outside of the brain, because this cortex lies on either side of the calcarine
fissure. This fissure, however, is clearly visible in a sagittal section
made between the two cerebral hemispheres.
The
primary visual cortex, with
its distinctive cell architecture, also corresponds to Area 17
described by the anatomist Brodmann in the early 20th century (link to Tool module
from the sidebar to the left).
The primary visual cortex
sends a large proportion of its connections to the secondary visual cortex
(V2), which consists of Brodmann's areas 18 and 19.
Though most of the neurons in the secondary visual cortex have properties similar
to those of the neurons in the primary visual cortex, many others have the distinctive
trait of responding
to far more complex shapes.
The analysis of visual stimuli that
begins in V1 and V2 continues through two major cortical systems for processing
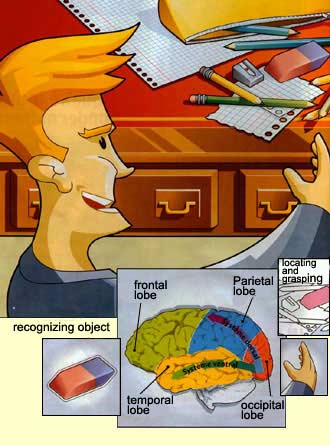
visual information. The first is the ventral
pathway, which extends to the temporal lobe and is thought to be
involved in recognizing objects. The second is the dorsal
pathway, which projects to the parietal lobe and appears to be essential
for locating objects.
Similarly to the other sensory systems
and the motor system, there is a correspondence or "mapping" between
the arrangement of the elements of the visual field as they strike the retina
and their arrangement on the surface of the visual cortex. This mapping onto the
visual cortex is called retinotopy, because it is the retina
that serves as the reference for the cortical maps of the various visual areas.
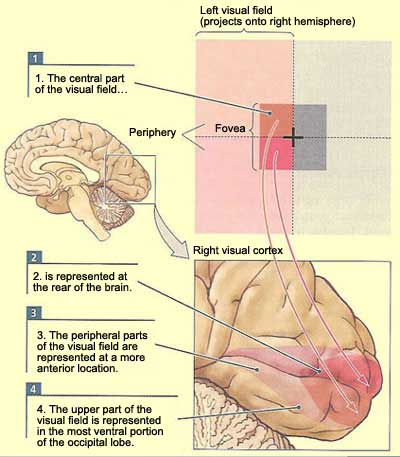
In retinotopic
maps, the zone of greatest discrimination in the retina—the fovea, a small
area at its centre—is represented by a disproportionately large area on the
cortex. The centre of the visual field, covered by the fovea, occupies the entire
posterior portion of the primary visual cortex, while the entire peripheral zone
of the visual field is analyzed in the remaining anterior portion.