Mitosis is
the type of cell division that takes place in human body
cells not only during development but also during tissue
renewal in adults. A cell that undergoes mitosis passes through
several stages that result in its division into two daughter
cells that resemble this mother cell. The daughter cells
may then divide in turn, which leads to the exponential growth
in the number of cells during embryonic development.
Meiosis is the other main type of cell
division. It occurs only in gametes (ova and spermatozoa).
Meiosis is specific to eukaryotic
cells and occurs when a diploid mother
cell produces four haploid daughter cells, each of which
has a different genome.
When fertilization occurs,
a male haploid cell fuses with a female haploid cell to create
a new diploid cell.
When a
stem cell divides, it can produce two new stem cells, or
two neuroblasts, or one stem cell and one neuroblast.
In the first case, the two daughter
stem cells will retain the property of producing other
pluripotent stem cells.
When a stem cell divides and at least one of its two daughter
cells is a neuroblast, differentiation is said to have
occurred. A differentiated cell is more specialized in
form and function than its mother cell. The differentiated
cell can produce only cells of its own particular lineage
and cannot go backward and produce stem cells.
The general
process that leads to the birth of new neurons is called neurogenesis.
Most of the neurons in the human neocortex are formed between
the fifth week and the fifth month of gestation.
Though most of the cortex’s neuronal development occurs
before birth, certain parts of the adult human brain retain
their ability to produce new neurons (follow the History Module
link below).
The process of neurogenesis in the adult brain is too limited
to replace any populations of neurons that might be destroyed
by injury or illness. But researchers hope that by learning
learn more about the processes that regulate this neurogenesis,
and in particular about the role
of environmental factors such as stress, they
may begin to devise ways of preserving this ability to grow
new neurons, as a treatment for degeneration due to pathology.
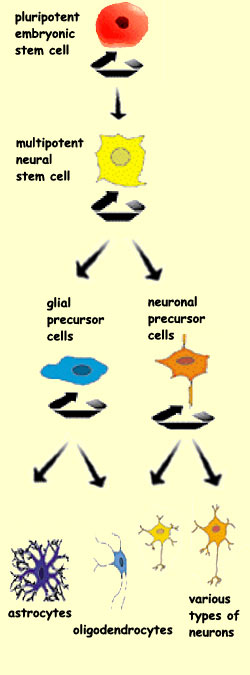
HOW STEM CELLS FORM NEURONS
In the space of just
a few months, all of a human being’s 100 billion neurons (with
a very few exceptions) and an even greater number of glial
cells are produced from a small population of precursor
cells.
The stems cells that proliferate in the ventricular zone
of the neural tube are the source of two major families
of cells in the nervous system: the neurons and the glial
cells. But the cell differentiation does not stop there. The
various structures of the brain are formed of countless
types of nerve cells that are distinguished by their neurotransmitters,
the molecules on the surface of their membranes, the types
of synapses that they form, and other such characteristics.
To generate all of this diversity, the processes
of cell proliferation, determination, and differentiation must
proceed in stages. At each stage, the ultimate destiny
of a cell is further defined. More specifically, this maturation
occurs through major changes in the replication and expression
of genes in the nuclei of these cells.
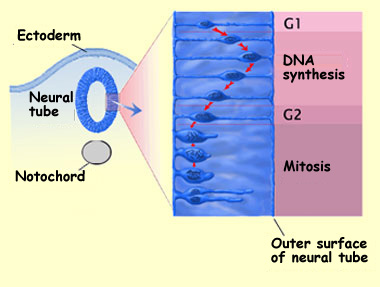
During the proliferation phase, for example, the cells divide
according to the usual cycle of mitosis (see sidebar), but the
cell divisions are accompanied by rather distinctive oscillating
movements of the cell nuclei. These nuclei go back and forth between
the ventricular zone of the neural tube (the part closer to its
central canal, or ventricle) and the marginal zone (the part closer
to its outer surface).
In phase G1, the growth phase, the newly divided cell begins by
extending a narrow cylinder of cytoplasm to the outer surface of
the neural tube, at the pia mater. The nucleus and the cytoplasm
surrounding it then move through this cylinder toward the outer
surface.
In phase S, the DNA-synthesis phase, the nucleus approaches the
outer surface and starts to replicate its DNA. Next comes phase
G2, the mitosis-preparation phase, in which the nucleus migrates
back toward the central canal of the neural tube while the cell
continues to develop. The cell then retracts its extension to the
outer surface and enters mitosis (phase M).
Based on material from Crump Institute
for Biological Imaging
Each
cell division gives rise either to more new stem cells or to
cells called neuroblasts that will differentiate into neurons
(see sidebar). The new stem cells will continue the mitotic
division cycle by sending out their own extensions to the outer
surface of the neural tube. But the neuroblasts will leave
the ventricular zone and migrate
to their final locations in the developing brain.
One possible explanation for this strange back-and-forth movement
might be that the nucleus needs to be exposed, in a particular
time sequence, to various cytoplasmic factors located in various
areas inside the cell. This phase of intense cell proliferation
results in an excess production of neurons, whose numbers will
then be reduced by apoptosis (for more on apoptosis, follow the
Tool Module link at the top of the left-hand column).
After their last mitosis in the ventricular zone, most of
the newly formed neuroblasts begin a migration that will
take them to their final position. Neuroblasts
guide themselves in various ways in the course of their migration.
In certain parts of the brain, such as the cortex and the
cerebellum, the migration of a large portion of these cells
is facilitated by radial glial cells that send out extensions
from the ventricular zone to the cortical surface.
The structure, function,
and activity of a cell depend in large part on its genes.
The sequence of cellular events that leads to the differentiation
of the neurons is thus controlled partly by intrinsic
factors—in other words, by cellular mechanisms
that activate and deactivate genes. For example, retinoic
acid, a derivative of vitamin A, activates specific receptors
that modulate the expression of certain genes.
But in eukaryotic
cells, the expression of the genes (or phenotype)
is always influenced as well by extrinsic factors (also
called epigenetic factors) from the cell’s environment.
Transplant experiments have thus
shown that if the grafted cells are taken from an animal
whose development is fairly advanced, these cells will
retain their original phenotype as they develop in their
new host. But the younger the animal from which the grafted
cells come, the more likely these cells will be to adopt
their host’s phenotype as they develop. This indicates
the important influence of the extracellular factors in
the new environment.
To understand the development of the
nervous system, we must thus constantly examine how the intrinsic
and extrinsic signals combine to ensure that the processes
of determination and differentiation take place successfully.
Some nerve cells,
such as motor
neurons, wait until they have arrived at their
final locations before sending out their axonal and dendritic
extensions. Other nerve cells, such as the granular
cells in the cerebellum, develop their extensions
while they are still migrating (see boxed text at the bottom
of this page).
HOW NEURONS CONNECT TO ONE ANOTHER
The location of each
neuron in the human brain is more critical than the locations
of the others cells in the human body, because neural functions
depend on precise connections between neurons and their
targets. In other words, the presynaptic and postsynaptic
components must be in the right place at the right time.
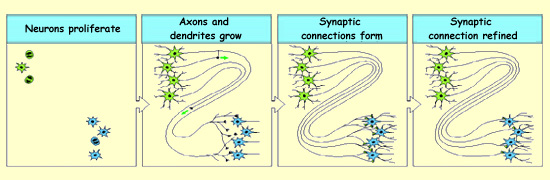
After the proliferation phase, the
neuron precursor cells leave the ventricular zone of the neural
tube and migrate to
their final locations in the brain. Once each neuron reaches its final destination,
its cell body develops
the axon and dendrites that will enable it to make connections with other
neurons.
To make their connections, the axons must find their specific target cells, which
is no small task. The process of forming an axonal pathway can be divided into
three phases: selecting the right route, choosing the right target, and finally
establishing a connection at the right destination.
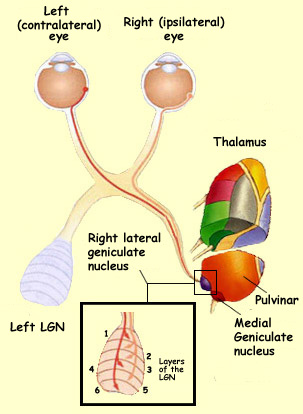
For example,
consider the case of an axon from a ganglion
cell in the retina that must reach the lateral
geniculate nucleus in the thalamus. Initially, this axon
follows the optic nerve, but very soon, it reaches the optic
chiasm, where it must choose one of three routes: the
left optic tract, the right optic tract, or the contralateral
optic nerve. Depending on whether this ganglion cell comes
from the nasal side or the temporal side of the retina, it
must select the correct route: either the
contralateral optic tract or the ipsilateral optic tract.
Even once it reaches the thalamus, the axon’s work
is not over. It still has a choice of a good dozen possible
targets. But it has to choose the right target on
the thalamus: in this case, the lateral geniculate nucleus
(LGN) and not the medial geniculate nucleus or the pulvinar.
Lastly, the axon must establish itself at the correct
final destination of its journey—in
this case, the correct layer of the lateral geniculate nucleus
and the proper retinotopic position.
Once it reaches its target, the axon develops a multitude of
synapses with it. A
selection process that depends on the activity of the neurons will
then reduce the number of these synapses so that only those that
play a significant role in the neural circuits are retained.
Source: Dr. Brian E. Staveley,
Department of Biology, Memorial University of Newfoundland
Long after birth, this mechanism of synaptic reinforcement associated
with neuronal activity continues to influence our synapses so
as to adjust our bodies to our activities and to our perceptions
of the outside world.
Some of the mechanisms employed in the early stages of embryonic
development will even adjust so that they continue to contribute
to the modifications that the brain undergoes throughout life.
This plasticity,
which enables us to adapt to the changing conditions in our environment,
is regarded as the basis
of human memory.
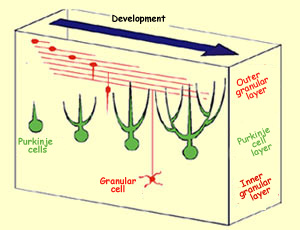
The first neurons
to be formed in the ventricular zone of the cerebellar
cortex are the Purkinje cells and
the Golgi cells, which migrate to the marginal zone immediately.
The ventricular zone also produces precursor neurons that
migrate beyond the Purkinje cell layer to form a second
germinal zone called the outer granular layer. It is from
this layer that the three main kinds of interneurons in
the cerebellum will develop: basket
cells, stellate cells, and granular cells. The
birth of these interneurons coincides with the elongation
of the dendrites of the Purkinje cells.
To reach their final destination,
the granular cells must migrate first through the
molecular layer and then through the Purkinje cell
layer. Upon arriving in the molecular layer, the
granular cells develop two extensions parallel
to the surface of the cerebellar cortex and perpendicular
to the dendrites of the Purkinje cells that are
in the process of developing. A third extension
soon forms and descends to the granular layer.
The body of the granular cell then simply has to
follow this extension to reach its final position
in the granular layer, leaving behind the two extensions
that form the parallel fibres which make connections
to the dendrite branches of the Purkinje cells.
Differentiation
of granular cells and Purkinje cells in the
cerebellum
Source: Mineko Kengaku, Laboratory for
Neural Cell Polarity