Each eye can be said actually to contain
not one retina but rather two retinas superimposed on each
other. One is composed of rods,
which are sensitive to the low levels of light that we experience
at dusk and dawn, for example. The other is composed of cones,
which can detect colour and are sensitive to broad daylight.
The retina is not designed to record the absolute intensity of the light reaching it, but rather to detect the differences in the intensity of the light striking it at different points.
THE RETINA
For you to see anything, your eye must
first form a precise image of it on your retina. Then the light energy
striking your retina must be converted into nerve impulses by the
retina's photoreceptor
cells.
The image can then be processed by your nervous system. This processing does
not start in the brain, but instead starts immediately in the retina itself.
In fact, anatomists regard the retina as a part of the brain that is located
outside it, somewhat the way you may regard your home satellite dish as an integral
part of your television receiver.
Physically, the retina is a thin layer of nerve tissue
with the consistency and thickness of a wet cigarette paper.
The neurons of the retina are arranged in 3
main layers separated by 2 intermediate layers whose
main purpose is to make connections among the various neurons.
The deepest layer of neurons processes the light first. These
neurons are the photoreceptors, the only
cells in the retina that can convert light into nerve impulses.
The photoreceptor layer then transmits these impulses to
the bipolar neurons in the second layer
and on to the ganglion neurons in the third
layer. It is only the axons of
these ganglion neurons that exit the eye and carry the nerve
impulses to the first
visual relay in the brain.
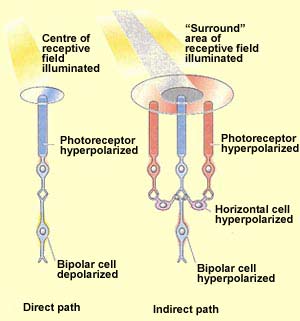
In addition to this direct pathway from the photoreceptors to
the brain, two other kinds of cells contribute to the processing
of visual information in the retina. The horizontal cells receive
information from the photoreceptors and transmit it to a number
of surrounding bipolar neurons. The amacrine cells receive
their inputs from the bipolar cells and do the same thing to the
ganglion neurons: activate the ones that are in their vicinity.
RECEPTIVE FIELDS, FROM THE RETINA
TO THE CORTEX
Each
of the neurons in the various
layers of the retina "covers" an area
in your field of vision. This area in space where the
presence of an appropriate stimulus will modify the
activity of this neuron is called the receptive
field of this neuron.
The receptive field of a single photoreceptor cell, for
example, can be said to be limited to the tiny spot of
light, within your field of vision, that corresponds
to this photoreceptor's precise location on your retina.
But in each succeeding layer of the retina, the receptive
fields become increasingly complex, and they become even
more complex when it comes to the neurons of the visual
cortex.
Here is an example of this complexity. The receptive
fields of bipolar cells are circular. But the centre
and the surrounding area of each circle work in opposite
ways: a ray of light that strikes the centre of the field
has the opposite effect from one that strikes the area
surrounding it (known as the "surround").
In fact, there are two types of bipolar cells, distinguished
by the way they respond to light on the centres of their
receptive fields. They are called ON-centre cells and
OFF-centre cells.
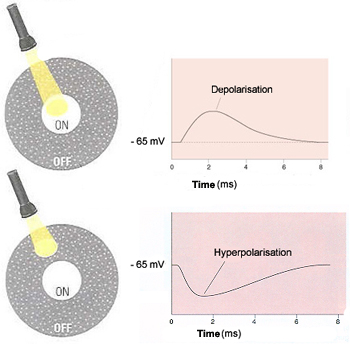
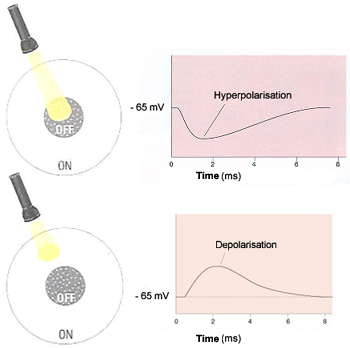
If a light stimulus applied to the centre of a bipolar
cells's receptive field has an excitatory
effect on that cell, causing it to become depolarized,
it is an ON-centre cell. A ray of light that falls
only on the surround, however, will have the opposite
effect on such a cell, inhibiting (hyperpolarizing)
it.
The other kind of bipolar cells, OFF-centre cells,
display exactly the reverse behaviour: light on the
field's centre has an inhibitory (hyperpolarizing)
effect, while light on the surround has an excitatory
(depolarizing ) effect.
ON-centre Bipolar Cell
OFF-centre Bipolar Cell
Receptive Field
of a Ganglion Cell
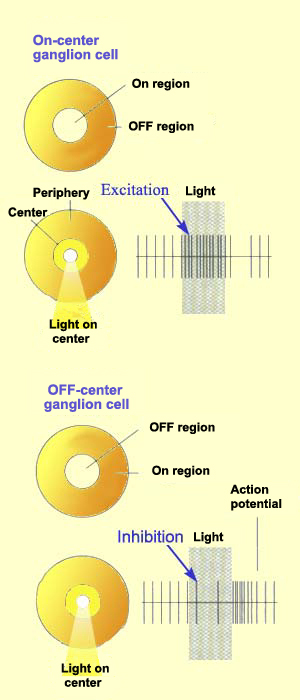
Just like bipolar cells, ganglion cells have concentric
receptive fields with a centre-surround antagonism.
But contrary to the two types of bipolar cells,
ON-centre ganglion cells and OFF-centre ganglion
cells do not respond by depolarizing or hyperpolarizing,
but rather by increasing or decreasing the frequency
with which they discharge action
potentials.
That said, the response to the stimulation of
the centre of the receptive field is always inhibited
by the stimulation of the surround.
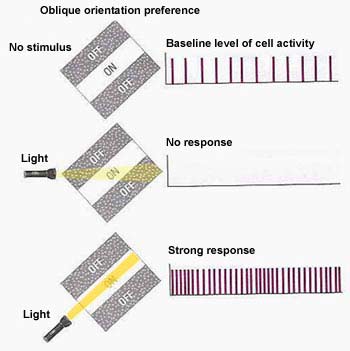
The receptive fields of the neurons
of the primary visual cortex are not circular,
but rectangular. They respond especially well
to rays of light that are oriented in a particular
direction. The cells whose receptive fields
thus respond to light with a specific orientation
are called simple cells.
These rectangular receptive fields often have
an ON centre band that responds positively to
light flanked by two OFF side bands that respond
to darkness. The diagram here shows that when
the beam of light is not oriented to follow the
ON band precisely, the stimulus is simply not
effective for this cell.
Simple Cell Receptive
Fields
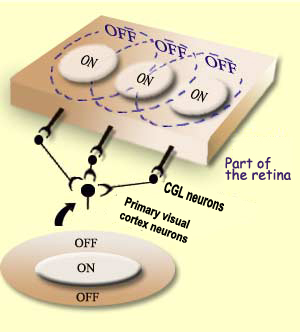
The simple cell receptive fields in the primary visual
cortex are thought to be the result of the convergence
of several adjacent receptive fields of cells in the relay
that precedes it, the lateral
geniculate nucleus. Note, by the way, that the receptive
fields of this nucleus are still circular, like those of
its source, the ganglion neurons in the retina.
The primary
visual cortex is the first relay in
the visual pathways where information from the two
eyes is combined. In other words, a single cell in
this cortex may respond just as much to the stimuli
presented to one eye as to those presented to the
other.
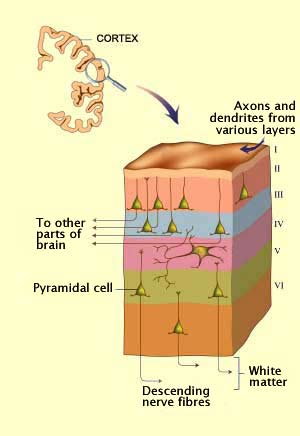
THE CELLULAR STRUCTURE OF THE VISUAL
CORTEX
In the visual cortex, the cell bodies of the neurons are
divided into six layers that typify the primate
neocortex. In this thin envelope of grey
matter, about 2 mm thick, the six layers are numbered
from I to VI, in Roman numerals, starting from the outside
(the layer in contact with the meninges).
Each layer is distinguished both by the type
of neurons that it contains and by the connections
that it makes with other areas of the brain.
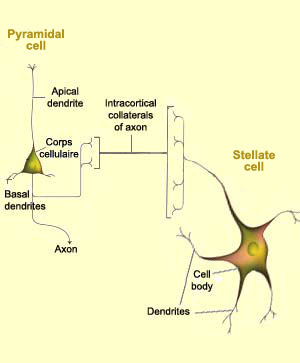
Layer IV, for example, contains numerous stellate
cells, small neurons with dendrites that radiate
out around the cell body and receive connections from the lateral
geniculate nucleus. Thus this layer specializes largely
in receiving information.
Pyramidal cells are found
in several layers of the visual cortex and are the only type
of neurons that project axons outside
it. Each pyramidal cell has one large dendrite, called the
apical dendrite, that branches upward into the higher layers
of the cortex, and other dendrites that emerge from the base
of the cell. Of course, each pyramidal cell also has an axon,
which may be very long to reach distant areas of the brain.
Layers III, V, and VI contain large numbers of pyramidal
cells and consequently serve as output pathways for the visual
cortex.
Layer I contains very few neurons.
It is composed of axons and dendrites from cells in the other
layers.