Even spinal reflexes, such as the
knee-jerk
reflex and the leg-withdrawal
reflex, which might appear simple at first glance, actually reveal all
the complexity of the spinal system for controlling movement. The activity of
the alpha motor neurons that innervate the muscles is subject to the triple influence
of sensory inputs, spinal interneurons, and the pyramidal tract.
For some time, the primary motor
cortex was thought to contain a detailed representation of every muscle in the
body, so that the activation of a particular pyramidal cell led to the activation
of a single group of motor neurons. But this view has been called into question
by recent studies suggesting that the pyramidal cells actually control groups
of muscles. The activation of these cells would therefore enable an entire limb
to be mobilized to accomplish a given action.
Moreover, recordings of
the activity of the motor cortex neurons during a body movement show that activation
begins before the movement and continues throughout its execution. This neural
activity may be encoding two main aspects of the movement: its force and its direction.
In addition to receiving
inputs from the premotor and supplementary motor areas, the pyramidal neurons
of the primary motor cortex receive information directly from somatosensory areas
3, 1, and 2. The other major source of incoming axons is the thalamus, or more
precisely the caudal part of the ventrolateral nucleus (designated VLc), which
relays information from the cerebellum.
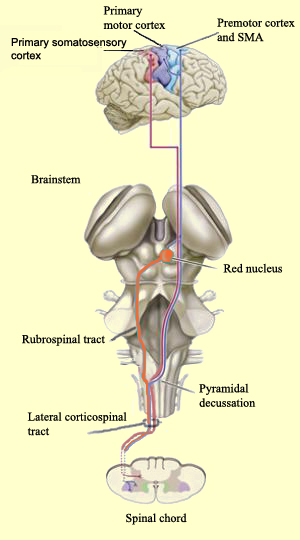
The primary motor cortex projects
its axons mainly to the corticospinal tract, which is composed of the lateral
and ventromedial systems.
The lateral system comprises two
main neural pathways, the larger of which is the lateral corticospinal
tract. Arising mainly in Areas 4 and 6 of the frontal lobe, which together
constitute the motor cortex, this is the longest neural pathway and one of the
largest in terms of the number of axons it contains (1 million). The other axons
in this tract arise mainly in the somatosensory areas and the parietal lobe.
After crossing the internal capsule, the midbrain, and the pons, the axons
arising in the cortex join together at the medulla oblongata to form a dense bundle
of nerve fibres. This bundle is shaped somewhat like a pyramid extending along
the ventral surface of the medulla, which is why it is called the pyramidal
tract at this location. (In contrast, all of the other tracts that arise
in the subcortical structures or the brainstem and follow a different route are
often referred to as the extrapyramidal tracts.)
Just
before entering the spinal cord, the pyramidal tract decussates. In other words,
the fibres from the left hemisphere of the cortex now cross over into the right
lateral column of the spinal cord, and vice versa. The axons of this tract ultimately
synapse on the motor neurons and interneurons of the dorsolateral portion of the
ventral horn of the spinal cord.
The rubrospinal tract
is the second tract in the lateral system. It arises from the neurons of the red
nucleus, in the midbrain. This nucleus receives information from the frontal cortex
, a region that also projects massively to the corticospinal tract. Indeed, over
the course of primate evolution, the role of the rubrospinal tract, an indirect
pathway, has diminished, while the corticospinal tract has assumed more and more
of the responsibility for motor control.
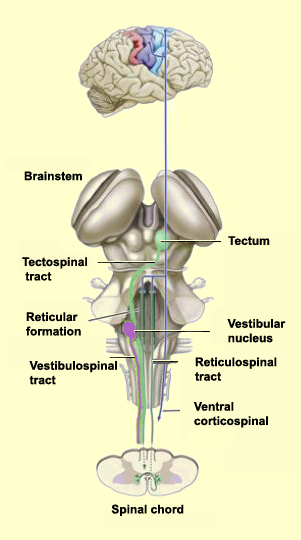
The
other major descending pathway, the ventromedial system, is composed
of four tracts that originate in various areas of the brainstem and contribute
chiefly to postural control and certain reflex movements. The originating neurons
of these tracts receive sensory information related to balance, body position,
and the visual environment.
The vestibulospinaltract
originates in the vestibular nuclei that receive information from the inner ear.
This tract helps to maintain the head in the correct position relative to the
shoulders, which is essential for continuing to look in a given direction while
the body is moving.
The tectospinal tract arises in
the superior colliculus (tectum) in the midbrain. The superior colliculus receives
some visual information directly from the retina, as well as somatosensory and
auditory information. Through the representation of the environment formed by
the superior colliculus, the tectospinal tract contributes to visual orientation.
The pontine (medial) and medullary (lateral) reticulospinal
tracts arise from the reticular formation nuclei in two main parts of
the brainstem: the pons and the medulla oblongata. The reticular formation receives
inputs from many sources and extends the entire length of the brainstem, from
the pons to the medulla. These two tracts help to maintain posture. The axons
originating in the pons enhance the spinal antigravity reflexes, while the axons
originating in the medulla have the opposite effect, releasing the muscles involved
in these reflexes and thus facilitating other movements.
In the mid-1990s, scientists studying
Area F5 in the ventral premotor cortex of monkeys found that certain neurons in
this area sent out action potentials not only when the monkeys were moving their
hands or mouths, but also when they were simply watching another animal or a human
being who was making such a gesture. These neurons were dubbed mirror
neurons because of the way that a visually observed
movement seemed to be reflected in the motor representation of the same movement
in the observer.
In addition to mirror neurons, which are activated both
when you perform an action yourself and when you see someone else performing it,
another kind of neurons, called canonical neurons, become activated when you merely
see an object that can be grasped by the prehensile movement of the hand whose
movements they encode—as if your brain were foreseeing a possible interaction
with this object and preparing itself accordingly.
What these two types
of neurons have in common is that they are both activated by an action regardless
of whether you are carrying that action out, anticipating carrying it out, or
watching someone else carrying it out. Because mirror neurons thus help us foresee
the consequences of our own actions, some have argued that these neurons may be
the cellular substrate for our ability also to understand the meaning of other
people's actions.
This understanding of other people's actions is the
foundation for all social relations, and especially for communication between
individuals. The discovery of mirror neurons may thus be particularly useful for
explaining how we can imagine
other people's intentions and state of mind. Lastly, the fact that Area F5
in monkeys is regarded as the homologue for Broca's area in humans suggests that
mirror neurons also are involved in human communication.
THE DISTINCTIVE CELL ARCHITECTURE OF THE
CEREBELLUM
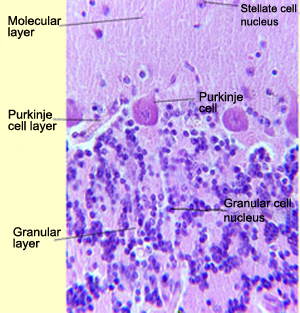
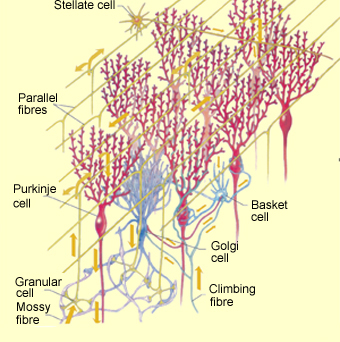
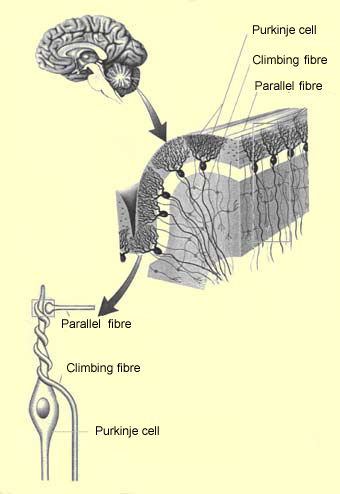
The Purkinje cells are
the most characteristic type of neurons in the cerebellum. The dendrites of each
Purkinje cell have a very distinctive pattern: their branches all lie in one plane
in which they assume the shape of a fan. The fan-shaped dendrites of adjacent
cells lie parallel to each other and are separated by a distance of about 0.1
mm. They deploy into the molecular
layer of the cerebellar cortex. The axons of the Purkinje cells synapse on
the neurons of the dentate nuclei of the cerebellum. These nuclei relay the information
to the thalamus, which then projects to the cortex and the striatum.
The dendrite branches of each Purkinje cell receive synapses from the branch
terminations of a single afferent climbing fibre.
This fibre is the axon of a neuron in the inferior olive, a nucleus in the medulla
oblongata. The inferior olive integrates the information from the muscle proprioceptors.
Each climbing fibre winds closely around the dendrites of its corresponding Purkinje
cell, so that the activation of this fibre will cause a massive excitation of
this cell.
In contrast, the second major source of inputs to the cerebellum,
the mossy fibres, act in a highly diffuse fashion. These fibres
are the axons of neurons in the pontine nuclei that receive information from the
cerebral cortex. The mossy fibres carry this information to synapses with the
small granular cells in the deep layer of the cerebellum. There
are so many of these granular cells that they are thought to account for half
of all the neurons in the brain!
The axons of these granular cells ascend
into the surface layer of the cerebellum (the cerebellar cortex) where they branch
into T shapes to form the parallel fibres. The parallel fibres
then run perpendicular to the Purkinje cell dendrite fans, thus crossing many
Purkinje cells and connecting them into a single contact. Though each parallel
fibre makes only one contact with each Purkinje cell that it crosses, it makes
contact with a huge number of such cells along its path, which measures just a
few millimetres. Likewise, each Purkinje cell receives over 100 000 synapses
from 100 000 different parallel fibres.
At first, this configuration
was believed to be the basis for the cerebellar
clock. Because the incoming message from the parallel fibres takes an increasing
amount of time to traverse the succeeding dendritic levels, a time lag develops.
It was thought that the cerebellum might use this lag to co-ordinate the sequence
of movements. But now the most commonly accepted interpretation of this dual afferent
system is that it provides the ideal basic structure for an elementary learning
mechanism called long-term
depression.
This depression occurs when the dendrite branches of
a Purkinje cell are activated by the climbing fibre and the parallel fibres simultaneously.
The result is a long-term reduction in the efficiency of the synapse between these
parallel fibres and the dendrites of this Purkinje cell.