Six oculomotor muscles are attached to each of your eyeballs and let you make various
types of movements with your eyes. Some of these movements
are slow, such as when your eyes follow an object passing
in front of them. Others, called saccades, are very rapid.
One good example of saccades is the movements that your
eyes are making while you are reading this page. Your eyes
stop for a moment at one point in a sentence to analyze
its image, then jump ahead very quickly to the next point.
THE EYE
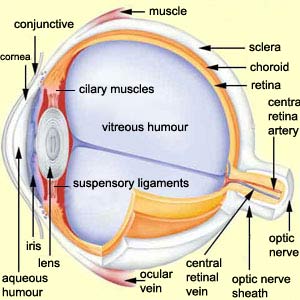
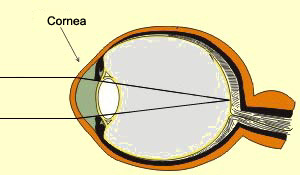
As can be seen in the cross-section
here, the first structure through which light passes into the eye
is the conjunctive—a thin, transparent
membrane that covers the front of the eye and then folds over
to line the inside of the eyelids.
Next, before light can reach
the various parts of the retina, it must
pass through the other optical components of the eye:
the cornea, the aqueous humour, the pupil (at the centre
of the iris), the lens, and, lastly, the vitreous humour,
a slightly more viscous fluid than the aqueous humour.
The cornea is continuous with the white of the eye, or sclera,
which forms the hard outer wall of the eyeball. Three pairs
of muscles are inserted into the sclera. It is these ocular
muscles that enable the eyeball to move in its
socket in the skull.
Between the sclera and the retina lies the choroid,
a richly vascularized layer that delivers nutrients to the
iris and the retina. The choroid contains many dark pigments
that make the inside of the eye, as seen through the pupil,
appear black
The aqueous humour and the vitreous humour play an essential role
in focusing the image on the retina. Because light travels at different
speeds in these two humours compared with its speed in the air,
they bend any light rays that enter the eye at a non-perpendicular
angle, so that these rays strike the proper place on the retina.
This process is called refraction.
The curvature of the cornea also accentuates
the refraction of the virtually parallel light rays that
reach the eye from very distant objects. Some of these rays
strike the cornea's centre. Hence they are already perpendicular
to the cornea, and their angle does not change. They continue
straight ahead to the centre of the retina. But other rays
from these same distant objects strike the curved parts of
the cornea. These rays are bent inward so that (provided
the person has no vision defects) they arrive at exactly
the same central point on the retina and form a focused image.
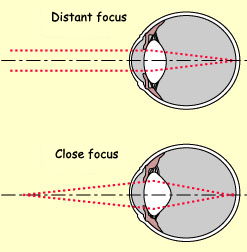
Where distant objects are concerned, the cornea does most
of the refracting of light rays to make them converge at
a single point on the retina; the lens also contributes,
but to a lesser extent. For closer objects, however (starting
about 9 metres from the eye), the lens plays a far more active
role in focusing the image on the retina. The light rays
reaching the eye from closer objects diverge more, so they
must be refracted more in order to converge on the retina.
The lens alters its own shape to provide this additional
refraction.
The lens is attached via the suspensory
ligaments to the ciliary muscles, which
in turn are attached to the sclera. These muscles form a ring around
the inside of the eye, so when they are relaxed, the tension on
the suspensory ligaments is greater. Consequently, the lens is flatter
and refracts less. But when the ciliary muscles contract, they relieve
the tension on the lens. It therefore tends to return to its natural,
more convex shape, which refracts light rays more, so that they
converge more. The increased refractive power that the lens thus
acquires allows a crisp image of close-up objects to be formed on
the retina. This phenomenon is called accommodation.
Even though the nerve
fibres that run from the left and right eyes to each
LGN connect to different layers within it, each of these
layers still maintains retinotopic mapping (as
is also found in the superior colliculus).
What is more, the maps in the six layers of each LGN
coincide, so that a line running through all of them
perpendicularly passes through neurons that are sensitive
to the same points in the binocular visual field. Hence
these points are perceived simultaneously by both eyes.
THE TARGETS OF THE OPTIC NERVE
The axons of the retina's
ganglion cells pass through the optic nerve, the optic chiasm, and the optic tractus.
They wrap around the midbrain and cross the medial surface
of the temporal lobe, and 80% of them then terminate in
a synaptic relay called the lateral geniculate
nucleus (LGN), located in the dorsal part of the
thalamus. The LGN is thus the major target for each optic
tract.
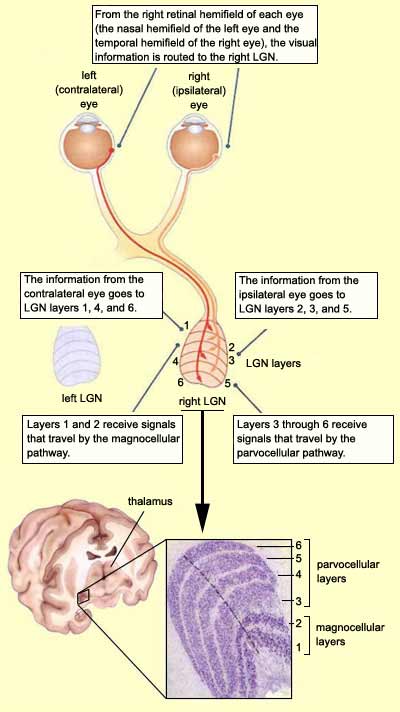
In cross-section, the left and right
LGNs are seen to be organized into six distinct cell layers.
When examined in three dimensions, these six layers look
like a stack of pancakes folded around the optic tract in
a way that resembles a knee joint, whence the name "geniculate",
from the Latin for knee.
The distribution of the LGN's neurons into various layers
suggests that some distinct aspects of the visual information
from the retina may be processed separately in this synaptic
relay. And that is exactly what has been demonstrated experimentally.
Because the right LGN processes all of the visual signals
from the left visual field (and vice versa for the left LGN),
it receives axons from the left nasal retina and the right
temporal retina. Research has shown that the axons from the
ipsilateral eye (in this case, the right eye) synapse on
cell layers 2, 3, and 5 in the LGN, whereas those from the
contralateral eye (here, the left one) project to layers
1, 4, and 6.
Examination reveals that the LGN's ventral layers (1 and 2) contain
larger neurons than its more dorsal layers (3, 4, 5, and 6). The
two ventral layers are therefore called magnocellular (M) layers,
while the four others are called parvocellular (P) layers,
just like the type M and type P ganglion cells. In fact,
it has been shown that it is precisely these type M ganglion cells
that project to the magnocellular layers of the LGN, and the type
P ganglion cells that project to its parvocellular layers. The
parallel processing in distinct information channels that begins
in the retina thus seems to be maintained through the LGN.
Even the small neurons that form the koniocellular layers on
the ventral side of each of the six numbered layers of the LGN receive connections
from the non-M non-P ganglion cells in the retina. This observation confirms
the segregation of information from different types of ganglion cells.
Despite the great influence that innervation from the retina has on the structure
of the LGN, about 80% of the excitatory inputs to the LGN come not from the retina
but from the primary visual cortex and lower centers in the brains! The primary
visual cortex thus appears to exert a significant feedback effect on the LGN.
In other words, the LGN's main target may in turn modify the LGN's own visual
responses.
Another observation lends weight to the idea that the LGN, just
like the other subcortical structures involved in vision,
does more than just passively relay information from the retina
to the cortex: the LGN may be activated by brainstem neurons whose
activity is associated with vigilance and with processes related
to attentiveness. These neurons seem to modulate the response of
the LGN neurons, which tends to confirm that the LGN is actually
the first location on the visual pathway where particular mental
states can influence our visual perception.
Unlike people who have colour blindness and confuse
certain colours, some very rare individuals cannot see
any colours at all. This disorder is called achromatopsia.
Achromatopsia can be inherited—for example, when there
is a genetic defect in the cones of the retina. But it
can also be acquired—as the result of a stroke, for
instance. Brain imaging studies in persons with achromatopsia
have confirmed the presence of substantial lesions in area
V4 of the visual cortex , which is known to be involved
in processing colours. These people describe their visual
perception of the world as dull and grey, something like
an old black and white television screen.
Achromatopsia is also usually accompanied by some difficulty
in recognizing the shapes of objects, which is consistent
with the shape-recognition role that also is played by
the ventral system.
THE
VARIOUS VISUAL CORTEXES
Hierarchical relationships
between a primary cortex and secondary cortexes are found
in several parts of the brain, including the sensory, motor, and auditory cortexes. The secondary
areas of these various cortexes then converge onto what
are called associative areas, which perform a more global
level of information processing. They progressively associate
signals from other sensory modalities to create an integrated,
multisensory representation of the world.
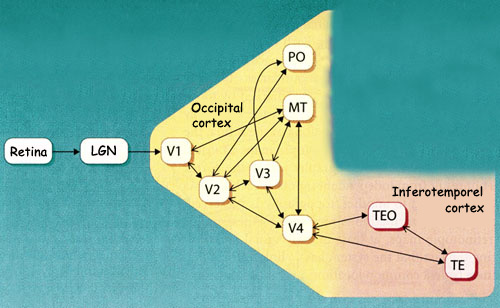
To date, researchers have discovered nearly
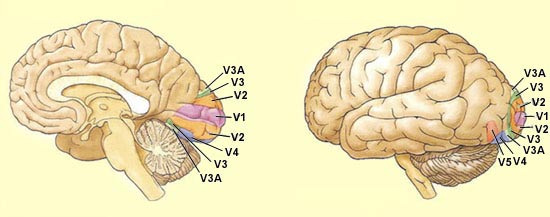
30 different cortical areas that contribute to visual perception.
The primary area (V1) and the secondary area (V2) are
surrounded by many other tertiary and associative visual areas:
V3, V4, V5 (or MT), PO, etc.
From all this complexity, however, a general
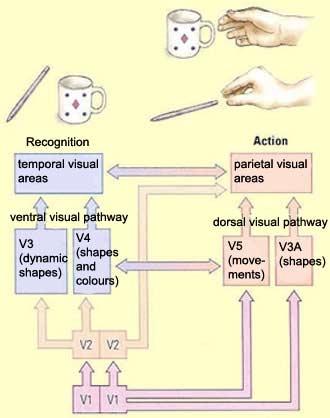
pattern does emerge. There appear to be two major cortical systems
for processing visual information: a ventral visual pathway that
extends to the temporal lobe, and a dorsal visual pathway that
projects to the parietal lobe.
The basic function of the ventral visual pathway seems
to be to let us consiously perceive, recognize, and
identify objects by processing their "intrinsic"
visual properties, such as shape and colour. The basic
function of the dorsal visual pathway seems to be to
let us exercise visual-motor control over objects by
processing their "extrinsic" properties—the
ones that are critical for handling them, such as their
size or their position and orientation in space.
In the cortical areas that contribute to the
ventral system, increasingly complex, specialized
representations of the outside world are elaborated.
Area V3 receives major
connections from area V2 and sends projections to areas MT and
V4. Many of the neurons in area V3 have properties similar to
those in V2. For example, most of them are selective for orientation.
But much remains unknown about area V3, and it also has some
cells with more complex properties. For example, some of these
cells are sensitive to colour and movement, traits more commonly
analyzed in subsequent stages of visual signal processing in
the brain.
After passing through areas V1, V2, and V3, part of the visual
information continues ventrally to area V4 on
its way to the temporal cortex. Area V4 receives information from
the blobs and interblobs of the striate cortex,
via a relay in V2. Like the cells in all of the other visual areas
besides V1 (also known as the "extrastriate areas"),
the cells in area V4 have larger receptive fields than those of
the striate cortex. Also, the receptive fields of V4 are often
sensitive to both colour and orientation. The exact role of area
V4 is still under debate, but it is probably involved in recognizing
shapes, and it appears to be essential for perceiving colours.
Area IT gets its name from the inferotemporal
cortex, where it is located, and comprises areas TEO and
TE. The cells of area IT receive many connections from
area V4 and respond to a very wide range of colours and
simple geometric shapes. These cells appear to play an
important role in visual memory, in addition to being a
key locus for object recognition.
Neurons have been
found in area IT that respond specifically to images
of faces. Initially discovered through intracellular
recordings in monkeys, the existence of these cells has
been confirmed in human beings through functional magnetic
resonance imaging (MRI) studies. This discovery is of
some interest to neuropsychologists, who have long known
of a rare syndrome called prosopagnosia, in
which patients have difficulty in recognizing faces,
even though the rest of their vision is normal. This
syndrome appears following lesions to the extrastriate
areas of the visual cortex, which strongly suggests that
these neurons in area IT may be involved.