In addition to using ordinary chemical
synapses to transmit nerve impulses through its circuits,
the retina also uses electrical synapses to transmit large volumes
of information more rapidly, especially in the pathways
that start in the rods. In addition, researchers have also
discovered that a great deal of neuromodulation takes place in the retina—diffusion
of substances over large distances to influence a large
number of neurons at once.
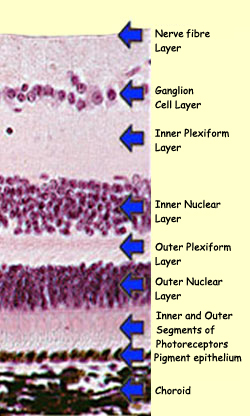
The ganglion cells are the last link
in the chain of neurons in the retina. This chain begins
in each retina's 125 million photoreceptors, which
gather information and channel it, via a small number of
synaptic connections, to the retina's 1 million ganglion
cells. The axons of these ganglion cells form the optic
nerve that carries the action potentials out of the eye
to the brain.
By the time these signals leave the eye, the information
that they carry is thus far more sophisticated than a mere
point-by-point representation of the world encoded by the
photoreceptors. And the retina is thus much more than a simple
layer of photosensitive cells. It is actually more like a
small brain outside the main one. In fact, in the development
of human embryos, the retinas are originally part of the
brain and detach from it subsequently.
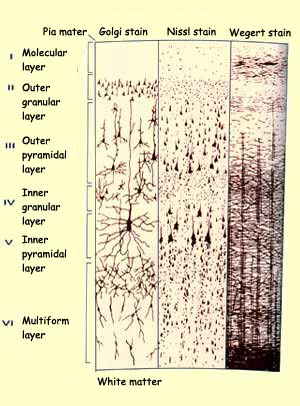
THE RETINA
Just like the rods and cones, whose structure and function are oriented entirely toward converting
light energy into nerve impulses, every other type of cell in the retina is
located and connected so as to perform some initial step in the processing
of visual information.
Source: University of Kansas Medical Center
While the other neurons in the retina emit only graduated
electrical potentials, the ganglion cells are
the only ones that send out neural signals in the form
of action
potentials. When you consider that it is the ganglion
cells' axons that form the optic nerve and thereby transmit
information from the retina over large distances, the significance
of the generation of action potentials in these cells becomes
apparent. Note that these potentials are generated spontaneously;
it is the frequency at which they are discharged that is
increased or decreased by the appearance of light in these
cells' receptive fields.
Though most ganglion cells have either ON-centre OFF-surround receptive
fields or the reverse, there are other criteria that
define other categories. On the basis of overall appearance,
neural connections, and electrophysiological traits,
at least three such categories of ganglion cells have
been distinguished in the retinas of macaques (short-tailed
monkeys whose retinas are very similar to our own).
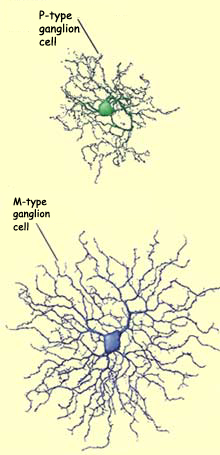
The small parvocellular (or "type
P") ganglion cells (from the Latin parvus, meaning "small")
represent about 90% of the total population of ganglion cells.
Large magnocellular (or "type M") ganglion cells (from
the Latin magnus, meaning "large") account
for about 5%. Non-M, non-P ganglion cells, which have not yet
been well characterized, account for the remaining 5%.
In addition to being larger
themselves, type M ganglion cells have larger
receptive fields, propagate action potentials more quickly
in the optic nerve, and are more sensitive to low-contrast
stimuli. In addition, the positive response of an M cell
to a stimulus consists of a brief salvo of action potentials,
whereas the response of P cells is more tonic, continuing
as long as the stimulus is active.
The most commonly accepted theory is that M cells are particularly
involved in detecting movement in a stimulus, whereas P cells,
with their small receptive fields, would be more sensitive
to its shape and details.
Another distinction is essential
for colour detection: most P cells and some non-M
non-P cells are sensitive to differences in the
wavelengths of light. Most P cells are in fact "single
colour opponent cells", which means that the response
to a given wavelength at the centre of their receptive fields
is inhibited by the response to another wavelength in the
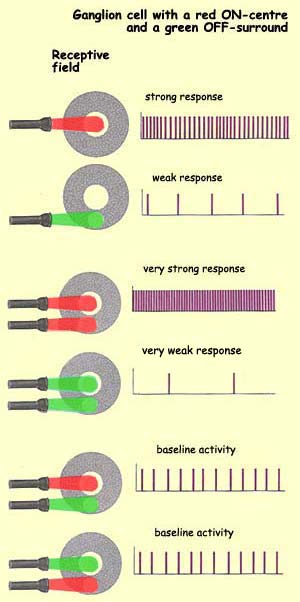
surround. In the case of a cell with a red ON-centre and
a green OFF-surround, red cones occupy the centre of the
field and green cones occupy the surround. The same thing
goes for cells with blue-yellow opposition, in which blue
cones are opposed to red and green ones. Type M ganglion
cells do not have any colour opposition, simply because both
the centre and the surround simultaneously receive information
from more than one type of cone. Also, there are no M cells
in the fovea, which confirms that these cells do not play
a role in processing colour.
Thus ganglion cells bring the brain information
that has already been partly processed as regards areal comparison
of the processes of light-dark, red-green, and blue-yellow opposition.
Like many other systems in the brain,
the visual system processes information in parallel.
Your two eyes first provide two parallel streams of information,
which your brain then compares to obtain an
estimate of the depth of a given object in the
scene in front of you.
The ganglion cells also appear to transmit independent
flows of information about the amount of light at each
point in space.
Type M ganglion cells can also detect subtle contrasts,
thanks to their very large receptive fields, while P cells,
with their small receptive fields, seem better adapted
to discriminating small details. Lastly, P cells and non-M
non-P cells are specialized in processing colours.
RECEPTIVE FIELDS,
FROM THE RETINA TO THE CORTEX
The centre-surround structure of the receptive
fields of retinal neurons results from the way that
the horizontal cells connect photoreceptors and bipolar
cells. This structure makes it possible to augment the
contrasts of objects in the visual field. Information
fed back from the inner plexiform layer also influences
the activity of the horizontal cells. These cells, in
return, modulate the signals from the photoreceptors
under various light conditions, thus making the process
of transduction less sensitive in bright
light and more sensitive in dim light.
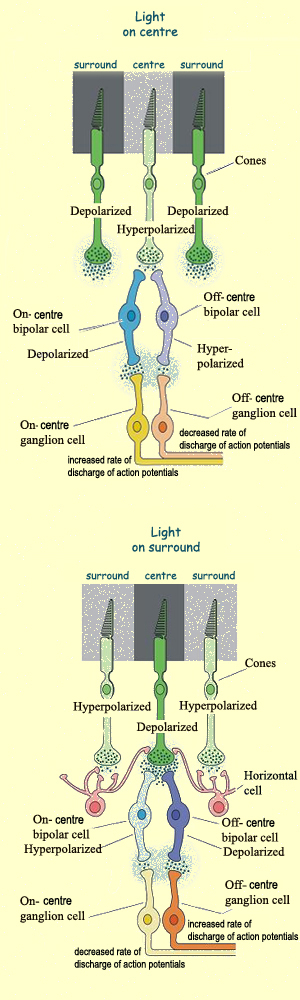
Bipolar cells, like all the other neurons in the retina
except the ganglion cells, transmit nerve impulses not
by means of action potentials, but in the form of simple
graduated potentials. Nevertheless, we speak of an ON response
when depolarization increases the amount of neurotransmitters
released and an OFF response when hyperpolarizaton reduces
this amount.
The question that then arises is whether the connections
between photoreceptors and bipolar cells are excitatory
or inhibitory.
In the absence of light, photoreceptors release their neurotransmitter,
glutamate, continuously. Consequently, the glutamate
receptors of OFF-centre bipolar cells are excitatory,
because the absence of light must stimulate them. Similarly,
the receptors of ON-centre bipolar cells are inhibitory, because
light striking the photoreceptors at the centre of their receptive
fields hyperpolarizes them and reduces their release of glutamate.
Since glutamate is an inhibitory neurotransmitter here (because
of certain metabotropic receptors), reducing it excites the
bipolar cell. Thus it is the excitatory or inhibitory nature
of the glutamate receptors that determines the type of receptive
field for bipolar cells.
ON-centre Ganglion Cells
Like bipolar cells,
ganglion cells have circular receptive fields, with centre-surround
opposition. In addition, the ON or OFF characteristic of a
bipolar cell is passed on to the ganglion cell to which it
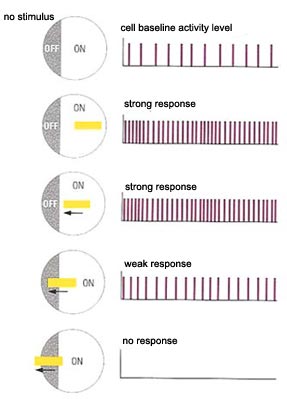
is connected. Most ganglion cells are not very sensitive to
light stimuli that strike both the centre and the surround
of their receptive fields. Hence, in total darkness or uniform
light, they emit few action potentials. However, these cells
are highly sensitive to differences in illumination at different
points in their receptive fields, such as when an area of light
or darkness sweeps across one side of a receptive field but
not the other.
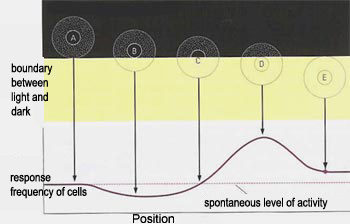
The information conveyed by the action potentials
from ganglion cells thus has more to do with the contrasts in illumination
between light and dark areas than with the absolute degree of luminosity. The perception of light and darkness therefore
is not absolute, but relative.
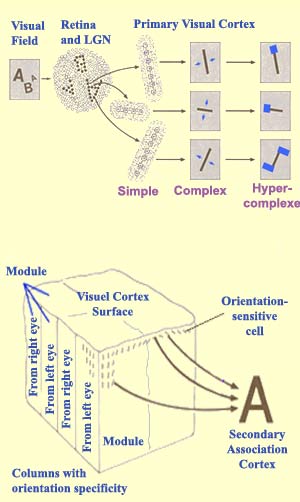
In the visual cortex, in addition to the simple and complex cells in the primary visual
area (V1, also known as Area 17 or the striate cortex) and
in secondary visual area 18 (V2), there are hypercomplex
cells in secondary visual area 19 (V5 or MT) that
respond only if a light stimulus presents a given ratio of
lit surface to dark surface, or is coming from a given angle,
or includes moving shapes. Some of these hypercomplex cells
also are sensitive only to lines of a certain length, so
that if the stimulus extends beyond this length, the cells'
response is reduced.
Hypercomplex cells occur when axons from several complex
cells with different orientations and adjacent visual fields
converge on a single neuron. These hypercomplex cells provide
yet another level of information processing. At every level,
each cell "sees" more than the cells at the levels
below it, and the highest-level cells have the greatest power
of abstraction. This capability is generated by the neuronal
connections at every stage along the visual pathways from
the eyes right up to the various visual cortexes in the brain.
These levels of abstraction can be summarized as follows:
the retina and the LGN "see" the position of an
object, the simple cells see its axis of orientation, the
complex cells see the movement of this axis, and the hypercomplex
cells see the object's edges and angles.
In addition to sending projections
outside the primary visual cortex, the axons of the pyramidal
cells in all of its layers also send out branches that
make local connections with one another. Most of these
connections are radial: they are made perpendicular to
the surface of the cortex and pass through its various
layers while remaining within the same column, thus preserving retinotopy.
However, the axons of certain pyramidal cells in layer III
send out branches that are horizontal rather than vertical
and hence make their connections across columns in layer
III.
These radial and horizontal connections play
distinct roles in the analysis of visual information.
In the visual systems
of newborn infants, the input pathways that convey information
from the two eyes to the brain converge on the same target
cells. But just a few weeks after birth, a segregation
occurs, and the connections are thenceforth made according
to which eye the input comes from. Following this synaptic
reorganization, each
layer of the lateral geniculate nucleus and each ocular
dominance column in the striate cortex receives inputs
from one eye only.
In order to study the effects of sensory deprivation during
critical periods of development, a number of experiments
have been conducted in which either one or both eyelids
of cats and monkeys have been sewn shut, or in which the
animals have been given strabismus surgically. These studies
have shown that the normal development of the connections
of the visual cortex depends not so much on
the activity of a particular neural pathway as on competition
between the relative activities of different pathways.
After the right eye of a young cat is sewn shut during
the critical period for the establishment of the ocular
dominance columns in the primary visual cortex, a process
of competition causes the surface area of the columns innervated
by the visual pathways of the sutured eye to decrease relative
to the corresponding area for the intact eye. This process
seems to work as follows. First, the axons projecting to
the cortex from the LGN cells that receive connections
from the closed eye regress, leaving neurons on the cortex
vacant. These neurons are then innervated by collateral
branches that develop from the axons of the cells of the
LGN of the intact eye.
THE CELLULAR
STRUCTURE OF THE VISUAL CORTEX
So much research has been done and published on the primary
visual cortex that we can now appreciate its cell
architecture in all its beauty and complexity.
First, there is the horizontal stratification of the
visual cortex into various types
of neurons that specialize in receiving or sending
neural information.
Next, the cortex is also divided radially, into a multitude
of columns in which all the neurons
respond to the same characteristic of a given point in
the visual field. The columns thus form functional units
that run perpendicular to the surface of the cortex.
In addition, if we insert
a microelectrode perpendicularly through the various layers
of the visual cortex, for example, all of the neurons that
it encounters will have the same orientation preference, regardless
of whether they have simple or complex receptive fields.
As a corollary, if we insert a microelectrode parallel to the
surface of the cortex, so that it passes through several columns
in the same layer, we will see the orientation preference change
as the microelectrode progresses. Hubel and Wiesel showed that
the orientation preference was reversed by 180 degrees on average
when the electrode moved about 1 millimetre in layer III.
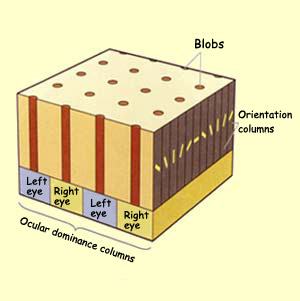
The ocular dominance
columns can be said to represent a third dimension
of the cell architecture of the primary visual cortex. These
columns are located in layer IV C and take the form of regularly
spaced bands 0.5 mm wide. In fact, experiments with
tracers (link to Experiment module from the sidebar to the
left) have shown that these bands represent the nerve endings
of the left and right eyes and that they thus alternate between
one eye and the other, in a pattern something like a zebra's
stripes.
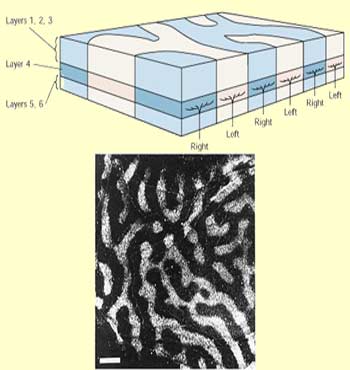
And as if all this were not
enough, in the late 1970s, other researchers, using a stain
called cytochrome oxydase, revealed the presence of another
kind of columns, spaced at regular intervals and running through
layers II, III, V, and VI. These columns, which look something
like a leopard's spots when viewed tangentially, are called blobs.
These blobs are arranged in lines and centred on an ocular
dominance band in layer IV C. Between the blobs are areas called
interblobs whose neurons do not have the characteristics of
these blobs.
What is special about the blob cells is that they are sensitive
to the wave length of light—in other words, its colour.
In addition, they are monocular, and they do not have any orientation
selectivity; instead, they have circularly symmetrical receptive
fields. Some blob cells even have the same centre-surround
colour opposition structure as the P ganglion cells where this
pathway originates (see box below).
Hubel and Wiesel also showed
that every point in the visual field produces a response
in a 2 mm x 2 mm area of the cortex. Such an area can contain
two complete groups of ocular dominance columns, 16 blobs
and interblobs that may contain more than two times all of
the orientations possible across 180 degrees. This region
of the cortex, which Hubel and Wiesel called a hypercolumn
(or, more generally, a cortical module)
seems both necessary and sufficient for analyzing the image
of a point in visual space. Because the cortex is a continuous
cellular layer and because it is very hard to establish the
boundaries of these modules physically, their existence from
a functional standpoint is still the subject of debate.
In the early 1960s, David Hubel and
Torsten Wiesel (who won the Nobel Prize for Medicine in 1981)
were the first to use microelectrodes to explore the receptive
fields of the neurons in the lateral geniculate nucleus and
the visual cortex. First, Hubel and Wiesel showed that the
neurons of the lateral geniculate nucleus behave practically
the same way as the ganglion cells in the retina. Then the
scientists discovered the existence of three relatively independent
pathways in the processing of visual information, each of
which takes care of a different aspect of vision.
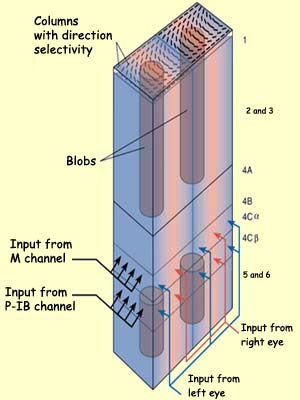
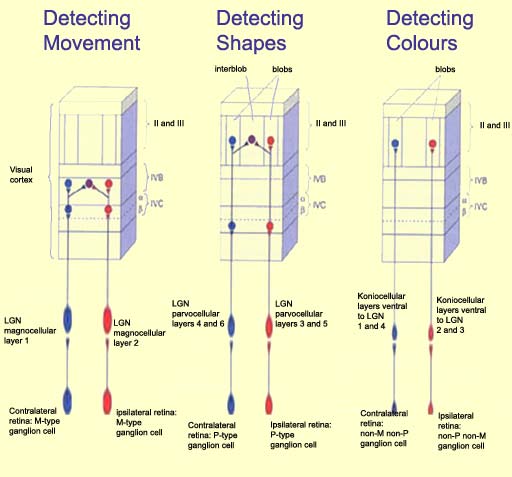
The first is the M (magnocellular) channel,
which begins in the magnocellular
ganglion cells of the retina, passes through the lateral geniculate nucleus, and projects
into layer IV
Caof
the striate cortex. In this layer, the receptive fields of
the neurons in this pathway are
no longer circular (as they are in the retina and LGN), but instead are somewhat oblong.
The cells in layer IV
Caproject
to the neurons of layer IV B. These latter neurons also have
simple receptive fields, but often respond to stimuli from
both eyes, contrary to the cells in layer IV C, whose receptive
fields are monocular. Thus the neurons in layer IV B begin
the process of integration that is necessary for human binocular
vision .
The cells in layer IV B are also selective for direction, but
only if the straight-line stimulus is moving in a particular
direction. For this reason, the M channel is thought to specialize
in the analyzing the movement of objects.
The second pathway for transmitting visual information is the P-IB
(parvocellular-interblob) channel, which starts with
the parvocellular ganglion cells in
the retina, passes through the LGN, and ends in the cells of
layer IV
Cb,
which respond like the parvocellular LGN cells from which they
arise: they have small, monocular, circular receptive fields,
most of them displaying red-green colour opposition.
The axons from the cells in layer IV
Cb then
project to the interblob areas in layer III. The complex cells in that layer are
even more specific than simple cells with regard to the orientation
of stimuli, which suggests that the P-IB channel specializes
in analyzing the shapes of objects.
The third pathway for transmitting visual signals is the blob
channel, which passes through the parvocellular and
koniocellular layers of the LGN and then converges on the blobs
in layer III. Besides certain neurons in layer IV C, the blobs
contain the only colour-sensitive neurons in the striate cortex.
The blobs are thus probably used to analyze the colours of
objects.