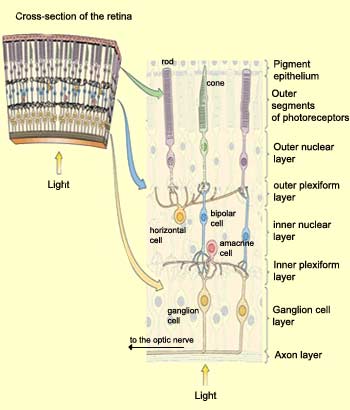
Intuitively, you might expect the photoreceptors to be located in the layer of the retina closest to its inner surface, so that they would receive the incoming light first. But that is not the case. Before reaching the photoreceptors, light must travel through all the other layers of the retina.
The reason for this somewhat paradoxical arrangement is that the light-sensitive pigments in the
photoreceptors must be in contact with the layer of epithelial cells at the back of the eye, which provide them with a continuous supply of retinene,
a light-sensitive derivative of Vitamin A. Also, after the structural arrangement of the retinene molecules has been changed by the light energy, they are recycled in this epithelium. The dark pigmentation of this epithelium also prevents unabsorbed photons from being reflected back onto the photoreceptors and thus creating light interference that would degrade the image.
Horizontal cells share a special characteristic with amacrine cells: the lack of any extension resembling an axon. These cells in fact have only dendrites, some of which are presynaptic, that is, play the role of axons. The extensions of these cells thus apparently play both roles.
THE RETINA
Within the retina,
information travels from the photoreceptors to the bipolar
cells and then on to the ganglion cells. At each stage along this
most direct visual pathway, the responses are modified by the
activation of lateral connections involving horizontal and amacrine
cells. Thus the analysis of visual stimuli begins even in the retina.
Curiously, in order to reach
the photoreceptors, incoming light must first pass through
all the other layers of cells in the retina (see sidebar).
The first of these is the ganglion cell layer,
composed of the bodies of ganglion cells. Next comes the inner
plexiform layer, a network of axons and dendrites
from ganglion cells, bipolar cells, and amacrine cells. After
that comes the inner nuclear layer, composed
of the bodies of bipolar, horizontal, and amacrine cells. Next
come the outer plexiform layer, composed of
the nerve endings of bipolar cells, horizontal cells, and photoreceptor
cells, and then the outer nuclear layer, which
contains the bodies of the photoreceptor cells. Last comes
the outer segment layer, containing the photoreceptors'
outer segments, in which the light-sensitive
pigments are located. The endings of these outer segments
are embedded in the pigment epithelium.
Every type of cell in these layers has its own distinctive distribution
pattern and physiological characteristics.
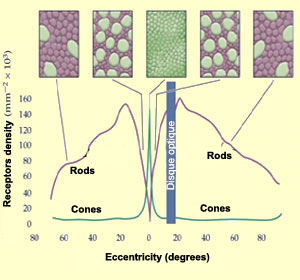
The distribution of the rods
and conesin the retina is not
uniform. Around the periphery of the retina, rods are far
more numerous, whereas in the centre, at the fovea,
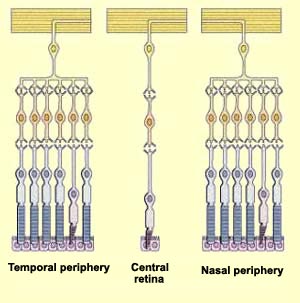
cones are. The number of photoreceptors connected to a single
ganglion cell is also far greater in the peripheral retina.
The combined effect of these distributions is to make the
peripheral retina more sensitive to light. The tradeoff is
that the convergence of many photoreceptors onto a single
ganglion cell causes the sharpness of the image to suffer.
For the high visual acuity found in
the central retina, a low ratio of photoreceptors to ganglion
cells is required. This acuity is also enhanced by the cones
in the fovea, which are very small and packed tightly against
one another. The farther from the fovea, the larger the cones
and the greater the space between them; the rods fill in the
remaining space. Despite the high density of cones in the fovea,
this region is so small that it contains only 1% of all the
cones in the retina.
Regardless of whether nerve impulses from the photoreceptor cells
are following the retina's direct pathway or its indirect pathway,
in which the horizontal cells are involved, these signals must
pass through the bipolar cells to reach the ganglion
cells. These signals are transmitted to the bipolar cells in the
form of graduated potentials, which may be either depolarizations
or hyperpolarizations, depending on whether the bipolar cell is
of the ON
or OFF type.
Each bipolar cell receives
some of its synaptic connections directly from a number of
photoreceptors located roughly opposite it. This number ranges
from one photoreceptor at the centre of the fovea to thousands
in the peripheral retina.
In addition to these direct connections from photoreceptors,
each bipolar cell receives some of its afferences from horizontal
cells. These cells are in turn connected to a set of photoreceptors
that surround the central group which connect to the bipolar
cell directly. As a result, the
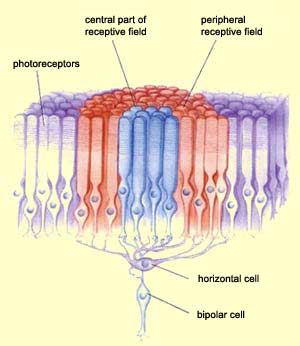
receptive field of a bipolar cell has two components: a
central receptive field composed of the information that travels
directly from the photoreceptors to the bipolar cell, and a
peripheral receptive field composed of information that arrives
via the horizontal cells.
Because the horizontal cells are connected laterally
to many rods, cones, and bipolar cells, their role is to inhibit
the activity of neighbouring cells. This selective suppression
of certain nerve signals is called lateral inhibition, and its
overall purpose is to increase the acuity of sensory signals. In
the case of vision, when light reaches the retina, it may illuminate
some photoreceptors brightly and others much less so. By suppressing
the signal from these less illuminated photoreceptors, the horizontal
cells ensure that only the signal from the well lit photoreceptors
reaches the ganglion cells, thus improving the contrast and definition
of the visual stimulus.
The retina's amacrine cells have highly diverse morphologies
and employ an impressive number of neurotransmitters. Their cell bodies are all
located in the inner
nuclear layer, while their synaptic endings are located in the inner
plexiform layer. By connecting bipolar cells with ganglion cells, they provide
an alternative, indirect path between them. Amacrine cells appear to have many
functions, most of them as yet unknown.
The circuits formed by the amacrine
cells in the inner plexiform layer provide additional information
to the ganglion cells, possibly by further increasing the
centre-surround contrast generated by the horizontal cells.
Whether a given synapse between
a photoreceptor cell and a bipolar cell is excitatory or
inhibitory may depend either on the type of neurotransmitter
released by the photoreceptor or on the type of receptors
in the postsynaptic membrane of the bipolar cell. The possibility
that one photoreceptor can release two different neurotransmitters
is receiving less and less credence, and all indications
are that ON and OFF bipolar cells have different molecular
receptors.
RECEPTIVE FIELDS, FROM THE
RETINA TO THE CORTEX
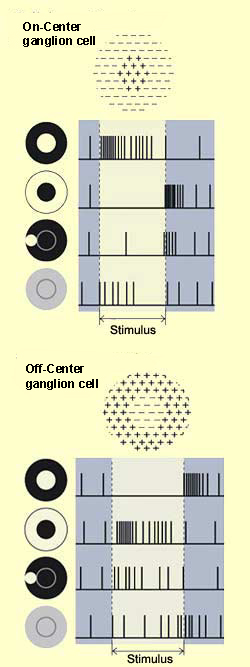
Bipolar cells have centre-surround receptive
fields. The centre of each such field receives direct connections
from a small number of photoreceptors, while the surrounding
area (called the "surround") receives inputs from a
larger set of photoreceptors whose activity is relayed by the horizontal
cells.
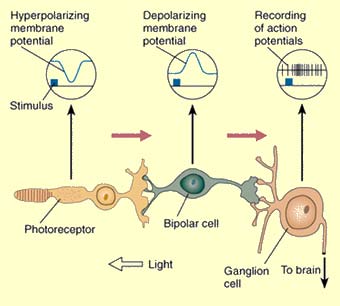
Light shining on the
centre of a bipolar cell's receptive field and light shining
on the surround produce opposite changes in the cell's
membrane potential. The diagram here uses an ON-centre
bipolar cell as an example.
If light is shined on the centre
of this cell's receptive field, the first change is a
hyperpolarization of the photoreceptor cell, causing
depolarization of the bipolar cell, because
of the inhibitory nature of the synapse between them.
This depolarization in turn excites the following cell,
a ganglion cell, causing it to emit action potentials
at a higher frequency.
Source: Adapted from J.E. Dowling
Conversely, if light were shined on
the surround of the receptive field of this same ON-centre bipolar
cell, it would become hyperpolarized. In contrast, another kind
of bipolar cell becomes depolarized when an area of darkness strikes
the centre of its receptive field, and hyperpolarized when it
strikes the surround. Bipolar cells of this kind are called OFF-centre
cells.
This centre-surround structure of the receptive fields
of bipolar cells is transmitted to the ganglion cells
via synapses located in the inner
plexiform layer .
Thus, some synapses connect ON-centre bipolar cells to
ON-centre ganglion cells, while others connect OFF-centre
bipolar cells to OFF-centre ganglion cells. The accentuation
of contrasts by the centre-surround receptive fields of
the bipolar cells is thereby preserved and passed on to
the ganglion cells, and ultimately to the visual cortex.
Human vision depends in large part on our ability to discern
contrasts between objects and the backgrounds behind them.
The establishment of parallel
pathways for the processing of visual information starting
in the retina is one of the mechanisms that makes this
discrimination possible.
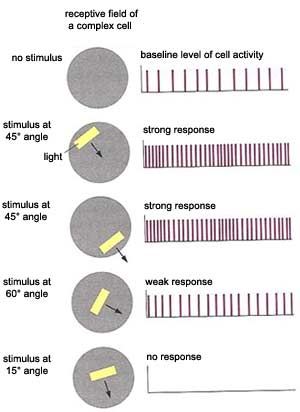
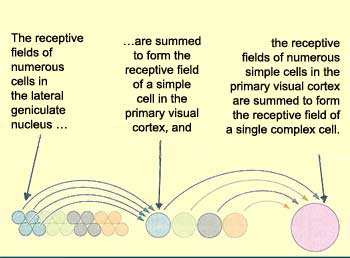
In addition to the simple
cells found mainly in layer IV of the visual cortex, there
are other cells, outside of layer IV, that respond to a light
stimulus only if it has a particular orientation and is moving.
These are called complex
cells. They detect movement through two mechanisms.
First, when the axons of many simple cells with the same
orientation and adjacent but not identical receptive fields
converge on a complex cell, it can detect movement from
the differences between these fields. Second, complex cells
can detect movement through the phenomenon of temporal
summation: if a cell that has already been excited once
is excited again shortly afterward, its membrane is still
depolarized enough that a stimulus that would not normally
suffice to trigger another action potential can do so.
Thus, when a moving light beam activates several simple
cells in succession, the temporal summation of the stimuli
applied to them causes the complex cell to respond to the
movement.
Complex cells also frequently display selectivity for direction,
responding only when the stimulus is moving in one direction
and not in the other. And unlike simple cells, complex cells
are not fussy about where the band of light is located in
their receptive field. Complex cells represent a further
level of visual information processing, but certainly not
the ultimate one, because researchers have also discovered
the existence of hypercomplex
cells.
Just as in the other relay points
in the visual pathways, there are far more cortical neurons
that receive information from the central part of the retina
than from its peripheral areas. This "retinotopy" reflects
a principle that operates in other parts of the cortex
as well: greater sensory or motor precision
requires the involvement of a greater cortical surface.
THE
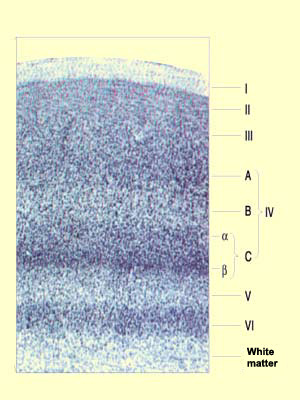
CELLULAR STRUCTURE OF THE VISUAL CORTEX
The primary visual cortex, like all the other parts
of the neocortex,
has a stratified cellular structure. Layers I to VI,
originally described by Brodmann, had to be further subdivided
as more was learned about the input and output pathways
of the visual cortex.
First, layer IV was divided into three sublayers designated
IV A, IV B, and IV C. Then layer IV C was itself subdivided
into IV Ca and IV Cb when
a difference was found between the connectivities of
the cells of the upper and lower parts of this sublayer.
The axons of the cells of the lateral geniculate nucleus transmit
information from the eye along various pathways that project
mainly into layer IV C. In addition, the neighbouring cells in
this layer receive receive information from neighbouring areas
of the retina, thus preserving a retinotopic structure. We also
know that the information flows emerging from the lateral geniculate
nucleus use
separate channels arising from its
internal structure.
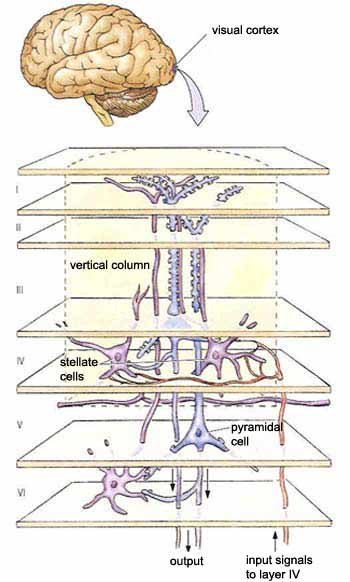
In layer IV C, these information streams are received
by the stellate
cells, whose axons pass them on to the dendrites of
the pyramidal cells in layers IV B and III. These pyramidal
cells then project their axons to other areas of the cortex.
As for the other output pathways from the primary visual
cortex, we know that the pyramidal cells in layer V project
to the superior colliculus and the pons at the subcortical
level, and that the axons from layer VI return massively
to the lateral geniculate nucleus, thus exerting a feedback
effect on this structure.
This stratification of the visual cortex
into horizontal layers can be readily revealed through simple
staining of its neurons. But the visual cortex is also organized
into vertical columns, which were not detected until electrophysiological
recordings were made of these neurons.
David Hubel and Torsten Wiesel were
the first scientists to propose this columnar structure superimposed
on the horizontal layers. Using microelectrodes to explore
the receptive fields of the neurons of the visual cortex,
they showed that this cortex can be regarded as a collection
of essentially identical columns. The difference from one
column to the next comes simply from the portion of the visual
field that is assigned to each of them. The succession of
functions of the various layers from the top of the column
to the bottom remains the same, but each
column processes a characteristic (contrast, colour, orientation,
movement, etc.) of a different part of the visual field.