The two kinds of photoreceptors in the
retinas of vertebrates—the rods and cones—differ
in many ways, both anatomically and functionally. The main difference
is the opposite roles that they play in vision. The rods provide
what is called scotopic vision: they are very sensitive to low levels
of light but cannot distinguish colours. The cones provide photopic
vision: they require bright light but let us see the world around
us in colour, and more sharply.
In both cases, however, the neural response is the same—the hyperpolarization
of the photoreceptor cells—and is initiated by the
same phenomenon: the absorption of light energy by photopigments
embedded in the discs of the photoreceptors' outer segments.
In the rods, the photosensitive pigment is rhodopsin, which
has its peak sensitivity at around 500 nanometres (nm) in the
visible-light band of the electromagnetic spectrum.
In the cones,
the photosensitive pigment is opsin, a
transmembrane protein that is very similar to rhodopsin.
Opsin comes in three different varieties, distinguished by
differences in their amino acid sequences that result in
differences in their light-absorption curves, with peaks
in the blue, green, and red portions of the visible light
spectrum, respectively.
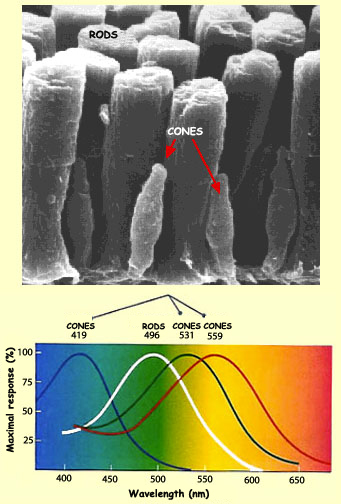
All three varieties of opsin are present in all cones. But
there are three types of cones, in each of which a different
variety of opsin heavily predominates, making it more sensitive
to a different part of the colour spectrum, as shown in the
diagram here. "Blue"
cones containing mostly blue-sensitive opsin are excited
chiefly by a wavelength of around 420 nm, "green"
cones by a wavelength around 530 nm, and "red"
cones by a wavelength near 560 nm.
But the attachment of color names to cones
can be misleading because the cones are not maximally sensitive exactly
in the red, green and blue parts of the spectrum. As you can see
on the picture above, the blue cones are most sensitive in the violet,
and the red cones in the yellow-green part of the spectrum. Consequently,
it is more accurate to refer to the three types of cones containing
mostly blue-, green-, and red-sensitive pigment as S-cones, M-cones,
and L-cones, respectively, where S, M, and L stand for short, medium,
and long wavelength.
An object whose colour falls anywhere in the visible spectrum will therefore
excite all three types of cones to varying extents. For example, a green object
will stimulate green cones for the most part, but also red cones to a lesser
extent, and blue cones to a still lesser one. Our perception of colours thus
depends on this superimposition of the various absorption spectra of the three
types of cones, and subsequently, of course, on the complex
neuronal interactions between the retina and the rest of the brain.
Colour blindness,
or daltonism, is a vision defect characterized
by the inability to differentiate certain colours or hues.
Its name comes from that of the English physicist John Dalton
(1766-1844), who suffered from this condition himself.
About 8% of all men are colour blind to varying degrees, and
slightly less than 1% of all women. The reason for this difference
is that the main form of colour blindness is hereditary, and
the genetic mutations that cause it occur on the X chromosome.
Since the mutated gene is recessive, women, who have two X
chromosomes, can carry the gene without being colour blind,
if the other X chromosome is unaffected. But men have only
one X chromosome, so if the mutated gene is present on it,
they are automatically colour blind.
Cases of total colour blindness, known as achromatopsy,
in which someone sees the world only in shades of grey, are
very rare. Usually, people who are colour blind have trouble
in telling red from green, or, much more rarely, blue from
yellow.
Classic red/green colour blindness is the result of a lack
of red cones in the retina. Forms of colour blindness are usually
classified according to the type of cone affected. Thus there
are three kinds of colour blindness, corresponding to the three
kinds of cones. Blindness to green, due to deficiency of the
green pigment, is called deuteranopia, and is the most common
form.
Dark adaptation is a two-step process
in which the eyes make the transition from photopic (cone-based)
to scotopic (rod-based) vision. Once you have spent a certain
amount of time in a well-lit room, your eyes' light-sensitivity
threshold is very high. If you then move into a darker room,
this threshold falls rapidly for the first 5 or 6 minutes,
then seems as if it were going to stabilize asymptotically.
But around the 7th minute, the threshold starts to fall even
more. About half an hour later, it reaches a second asymptotic
level, much lower than the first. This minimum level is the
threshold for scotopic vision, whereas the initial level
represented the threshold for photopic vision.
HOW LIGHT GETS
CONVERTED INTO NERVE IMPULSES
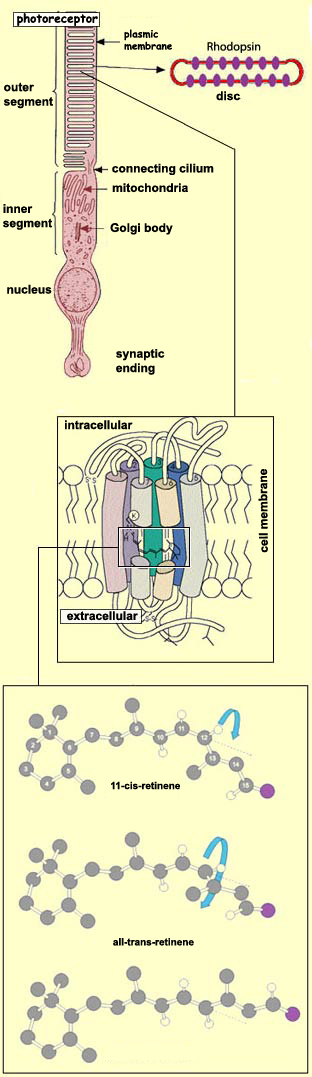
The transduction
of light energy into variations in photoreceptors' membrane
potential begins with the absorption of photons by light-sensitive
pigment proteins in the discs of the photoreceptors' outer
segments. These pigment proteins belong to a family known
as the opsins. The pigment protein in
rods is called rhodopsin, while the pigment
protein in cones is called iodopsin. A
single rod can contain up to 100 million molecules of rhodopsin
in its outer segment discs.
Rhodopsin molecules contain seven transmembrane domains
and somewhat resemble metabotropic
synaptic receptors in their structure. In fact, rhodopsin
can be regarded as a receptor protein whose agonist is
already bound to it. This agonist is a molecule called retinene (or
retinal) that is derived from Vitamin A. It is bound to
the middle of the seventh transmembrane domain of the rhodopsin
molecule. More specifically, it is the 11-cis form
of retinene, which, when it absorbs light, isomerizes to
the all-trans form. This isomerization converts
the rhodopsin to its active form, metarhodopsin II. This
reconfiguration of the retinene molecule thus produces
the same effect as if a neurotransmitter had suddenly bound
to a receptor.
Next, the metarhodopsin II stimulates a particular G protein
called transducin. Like
all G proteins, the transducin then activates another
enzyme, in this case phosphodiesterase (PDE). When the
PDE is activated, it converts the cGMP that is present
in the rod's cytoplasm in the absence of light into regular
GMP. The resulting decrease in cGMP closes the sodium channels
in the rod's cell membrane, thus hyperpolarizing the cell.
As a result, fewer neurotransmitter (most likely glutamate)
molecules are released from the photoreceptor's synaptic
endings.
Note that a signal amplification occurs at two points in
this biochemical cascade. Every metarhodopsin II molecule
activates about 100 G-protein molecules, and every phosphodiesterase
molecule hydrolyzes about 1 000 molecules of cGMP into
GMP. It is this amplification phenomenon that enables rods
to detect the presence of a single photon of light.
The Vitamin A that our bodies produce
from the beta carotene in many of the foods we eat (including,
most famously, carrots) is needed to synthesize the retinene
bound to the centre of the rhodopsin molecule. Indeed, a severe
Vitamin A deficiency impairs night vision, because of the smaller
amounts of retinene being produced. During the daytime, however,
there is generally enough light to allow relatively normal
vision despite low levels of visual pigments.