Another process that
helps axons to find their targets is fasciculation, whereby
the neurons that develop together intertwine and form bundles.
Because the surface area of the extracellular matrix is limited,
this enables several neurons to literally pile on top of
the one that is advancing across the substrate. Fasciculation
is made possible by the expression of molecules on the surface
of the neurons’ membranes, called cell
adhesion molecules (CAMs). The CAMs on
neighbouring axons interact so that they can adhere to one
another and grow toward their target as a group.
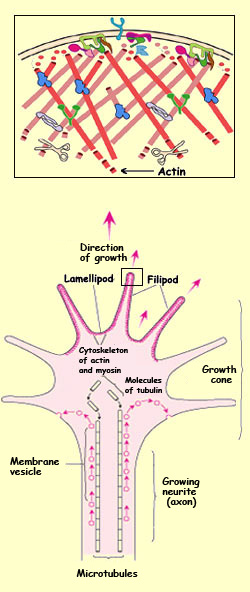
THE GROWTH CONE
The
growth cone is the ameboid structure that is located at the
tip of a growing axon and that pulls it forward. The growth
cone moves through a sequence of phases: protrusion, adhesion,
and contraction.
To protrude in the right direction, the growth cone depends
on complex cellular machinery that responds to molecular signals
received from the environment by specialized receptors. For
example, we know that it is the polymerization of actin that
causes the growth cone to advance, and that the biochemical
cascade that triggers this polymerization involves Rho GTPases.
The actin filaments in the growing axon project into the filipodia
of its growth cone. In a less organized fashion, these filaments
also spread through the lamellipodia, where they form a sort
of lattice. Near the periphery of the growth cone, the actin
filaments elongate by polymerization of the actin proteins.
In contrast, in the central part of the lamellipodia, the actin
filaments are broken down. (This same process is found in all
cells that move by protruding their lamellipodia.)
At the centre of the axon that elongates behind the advancing
growth cone, a cytoskeleton made of tubulin proteins
forms. Vesicles travel along these microtubules and fuse with
the growth cone’s membrane to let it expand.
Source: Max-Planck-Institut
The axon’s growth depends on the molecular
interactions between its growth cone and its substrate. One of
the main components of this substrate is the extracellular matrix,
composed of fibrous proteins in between the cells. Through this
matrix, axon growth paths are marked by permissive proteins such
as laminin. Laminin binds with molecules called integrins on
the growth cone membrane to encourage the growth cone to advance
in the right direction. The substrate also contains other, repellent
proteins that control the axon’s elongation even further
by preventing it from growing in certain other directions. Thus
the elongation of axons is controlled by molecular “direction
signs” all along the routes that they travel.
One of the great challenges
for the neurons in the developing human motor and sensory
nervous systems is to ensure that adjacent points in the
peripheral parts of these systems are represented by adjacent
points in the centres where their signals are processed in
the brain. The growing axons organize themselves with uncanny
accuracy, forming precise topographical maps such as the motor
homunculus and the somatosensory homunculus, and for
some time, researchers have been investigating how they do
so.
In the early 1960s, following experiments on the visual pathways
of frogs, researchers hypothesized that there was a chemical
affinity between molecules on the growth cones of the various
axons and other molecules on the target cells that these axons
had to reach. In other words, it was somewhat as if each axon
from the peripheral nervous system received a chemical label
that told it which position to head for, while a complementary
label on the target cell enabled each axon to find and reach
its assigned position.
But this hypothesis of a “key-in-lock” type of
correspondence, where there would have to be just as many different
types of keys and locks as there were different positions,
was contradicted by the experimental data. In fact, other experiments
have how shown that there is indeed a correspondence between
some molecules on the growth cone and others in the target
structure, but this correspondence is achieved by a concentration
gradient that uses only one set of keys and locks.
Source:
Dr. Brian E. Staveley
Department of Biology
Memorial University of Newfoundland
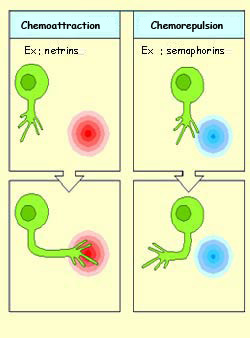
Among those diffusible
molecules that guide the growth cone by chemoattraction, the
group that has been best characterized is the netrins.
These molecules are secreted by the target cells, diffuse
into the extracellular environment, and influence the growth
cones of certain axons over long distances. Netrins resemble
the CAMs such as laminin that are found in the extracellular
matrix and that guide the growth cone by direct contact.
Like the laminins, the netrins influence the growth cone
by binding to specific transmembrane receptors that transduce
their signals.
In the developing embryo, netrins are often secreted near
a point where axons have to choose whether to stay on the
same side of the midline or go across it. For example, we
now have good descriptions of the role of netrins in the
development of the spinothalamic tract, which relays information
about heat and pain stimuli from the periphery to the thalamus.
Netrins also help some axons of the central nervous system
to cross the midline (for instance, in the corpus callosum
and the optic
chiasm).
Molecules that guide the growth cone by chemorepulsion can
be either non-diffusible (bound to the cell membrane) or
diffusible (secreted by the cell and diffused into the extracellular
environment). Semaphorins are one family
of molecules that guide the axons’ growth cones by
repelling them.
The molecules that guide axon growth are
not attractive or repellent in and of themselves. Instead, they
acquire one or the other of these characteristics depending on
the type of receptor expressed by a particular growth cone. Just
as in synaptic transmission and many other processes that trigger
cascades of biochemical reactions, it is the electrochemical
affinity between a molecule and its receptor that sets in motion
the internal mechanisms that will cause the growth cone to change.
Thus, the well known guidance molecule netrin-1 attracts
the interneurons of the dorsal commissure but repels certain
classes of motor neurons. A given guidance factor can thus
be interpreted in two completely different ways by the
growth cone, depending on what type of receptors it expresses
on its surface.
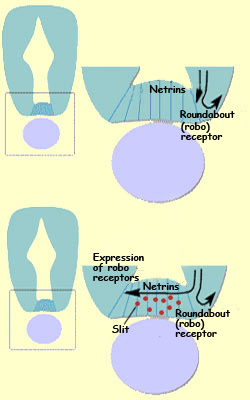
To continue with our example, once the netrins have attracted
the interneurons’ growth cones into the medial ventral
portion of the spinal
cord, these growth cones begin to express more roundaboutreceptors (or robos).
These receptors are sensitive to a repellent factor called Slit that
is also secreted in the medial ventral region of the spinal
cord. Consequently, the growth cones then begin to turn
away from this region, which causes them to continue across
the midline.
Section of the neural tube with
an enlargement of the floor plate (the lavender oval represents
the notochord)
In this example, the cells on the midline
of the spinal cord that secrete these various guidance molecules
act as intermediate targets, initially attracting the growing
axons, then repelling them toward their final targets.
NGF is produced by the
target cells of the sympathetic
nervous system. If this system is neutralized by
the injection of specific antibodies, the sympathetic ganglia
degenerate completely. The same thing happens if axoplasmic
transport is interrupted: the neurons die despite the release
of NGF by the target tissue.
NGF is
a protein that contains three sub-units, designated alpha,
beta, and gamma. The gamma sub-unit has proteolytic enzymatic
properties. The alpha sub-unit inhibits the protease. The
beta sub-unit is responsible for biological activity.
NGF resembles other growth-promoting
proteins (such as insulin), which suggests that they come
from the same family of molecules. There are many other neuron
growth factors; for example, motor neurons do not respond
to NGF.
TROPHIC FACTORS
AND NEURONAL DEATH
Over a long period that begins at
birth and lasts until adolescence, the number of neurons and
synapses that each individual is born with decreases quite
substantially. This neuronal death is the result of a competition
for trophic factors that are secreted in limited amounts by
the neurons’ target cells.
The first of these trophic factors to be discovered is a peptide
called nerve growth factor (NGF). It was discovered
in the early 1950s by Italian developmental biologist Rita Levi-Montalcini.
She was awarded the Nobel Prize in Physiology or Medicine for
this work jointly with American biochemist Stanley Cohen in 1986.
NGF is recognized by the sympathetic axons, which transport it
back up to their cell bodies, where its molecular interactions
promote these neurons’ survival.
The way that NGF and other
neurotrophins promote survival is by blocking a genetic program
contained in each cell that would otherwise cause it to self-destruct,
a process called apoptosis (for more on apoptosis,
follow the Tool Module link to the left). Unlike necrosis, an
accidental process of cell death that causes inflammation, apoptosis
is a methodical disassembling of the cell’s components
that takes place gradually. This process of programmed cell death
allows an optimal configuration of the brain’s wiring by
a) selecting those cells whose connections have proven the most
efficient, b) balancing the numbers of presynaptic and postsynaptic
neurons, and c) eliminating neurons that are malformed or damaged.
Not all of the various neurotrophins act on the same neurons.
NGF, for example, enables the survival of the parasympathetic
neurons, while BDNF enables that of neurons in the sensory ganglia,
and NT-3 that of both of these neuronal populations. The selective
action of the neurotrophins is attributable to a family of receptors
called Trk receptors, thus designated because these proteins
were initially identified as receptors capable of activating
the enzyme tyrosine kinase.
Source: Dr. Paul Kelly
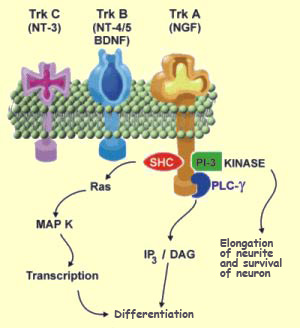
The family of Trk
receptors for neurotrophins includes: TrkA, which
is a receptor mainly for NGF; TrkB, which is a receptor for
BDNF; and TrkC, which is a receptor for NT-3. Given the close
structural similarities among the various neurotrophins on
the one hand and among the various Trk receptors on the other,
there is a certain amount of cross-activation among them. Thus,
under certain conditions, NT-3 can bind to and activate TrkB
receptors.
The positing
of an epigenetic form of evolution that takes place through
the selection of synapses has given rise to many debates
on the possibility that a sort of “neural Darwinism” somehow
continues the process of the genetic evolution of biological
species, but at the level of the individual and, consequently,
of the social group.
Some scientists, such as biologist
Richard Dawkins, have used the term “memes” to
designate these selected cognitive states that can be transmitted
from one individual to another. Memes would thus be a full-fledged
form of life whose substrate consisted of the human brain.
In the course of development,
long before any neuron growth factors are secreted, a large
number of molecular inductions occur. This
is what happens when one group of cells influences the development
of another by secreting morphogenic substances.
For example, the formation of the neural
plate from the ectoderm is induced by the cells of the
mesoderm.
These induction factors, most of which have some similarities
to growth factors, act by stimulating receptors in the cell
membrane, which influences the activity of certain genes involved
in cell differentiation.
Various induction factors can be secreted in succession to
induce different stages of differentiation. Or alternatively,
a single induction molecule can be diffused into its environment
to create a concentration gradient, just as chemotrophic
guidance factors do. In this case, exposure to differing
concentrations of the same substance can also result in differentiations
into distinct types of cells.
FORMATION AND
SELECTIVE STABILIZATION OF SYNAPSES
When the growth cone of an axon comes into
contact with its target cell, it flattens and forms a synapse with
this cell. When the axon is from a motor neuron and the target
cell is a muscle fibre, this synapse is called a neuromuscular
junction, and neuromuscular junctions are the kind of synapses
whose formation mechanism we know the most about.
We know, for example, that the proteins secreted by the motor
neuron’s growth cone help to establish receptors on the
membrane of the postsynaptic muscle fibre at the precise location
where the growth cone comes into contact with it. One of these
proteins is agrin. Agrin binds to the muscle-specific
receptor tyrosine kinase (MuSK), which is widely distributed
throughout the muscle cell membrane. The MuSK receptors to which
the agrin binds then activate an intracellular protein called rapsyn.
The rapsyn then causes nicotinic
cholinergic receptors to congregate directly below the growth
cone on the muscle cell membrane. (More recent studies have shown,
however, that the muscle cell may not need the growth cone to
initiate the task of congregating the receptors and can do so
on its own, thus helping to determine the location of its motor
end plate.)
At the same time, factors secreted by the target cell (in this
case, the muscle cell), will stimulate the entry of calcium ions
into the growth cone. This will have several effects on the presynaptic
ending, one of which will be to alter its cytoskeleton so as
to give the presynaptic button its flattened shape. The influx
of calcium ions will also promote the release of neurotransmitters.
A few weeks later in the embryo’s development, this rudimentary
form of synaptic
transmission will produce a synapse that is capable of transmitting
neural activity.
In the striate muscles of vertebrates,
cholinergic receptors are confined to the motor end plates, where
the density of receptors can be as much as 20 000 per square
micrometre, compared with scarcely 50 elsewhere on the muscle
fibres. If we sever the motor nerve that innervates a muscle
fibre and let this nerve degenerate, the receptors will become
more densely distributed throughout the muscle fibre but never
so densely as they originally were at the motor end plate. And
if the muscle becomes innervated again, the receptors that appeared
away from the motor end plate will disappear, leaving only those
inside this region.
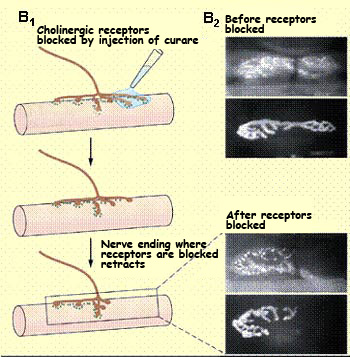
Experiments have also
shown that even when the acetylcholine receptors at the
neuromuscular junction are blocked with an antagonist such
as curare, polyneuronal innervation of a single muscle
fibre persists. Similarly, if the emission of action potentials
from the motor neurons is blocked with a sodium-channel
blocker such as tetrodotoxin, no reduction in innervation
occurs.
These experiments clearly show that the process by which
various axons compete with one another to connect to a
particular target cell is regulated by the electrical activity
of the presynaptic and postsynaptic partners.
Indeed, the subsequent stages of the synapse’s
development will also depend on neural activity. This activity
will significantly influence the competitive interactions among
neurons. These interactions do not actually involve a “winning” axon’s
crowding out “losing”ones. What happens instead is
that the specialized postsynaptic structures associated with
certain axons gradually disappear, thus reducing the electrical
activity between these axons and the muscle cells and further
encouraging the loss of receptors on these synapses. This process
would naturally cause the axons in question to retract and atrophy.
In parallel with the adjustment
of the size of the populations of presynaptic and postsynaptic
neurons, synaptic reorganization thus represents
the final step in establishing the circuits of the brain.
And contrary to the preceding steps, neuronal activity
and synaptic transmission thus play an essential role
in the fine-tuning of the neural networks.
The fact that mechanisms of competition and elimination
of synapses come into play in the course of development
had been known for several decades, but the idea that this
elimination could take place epigenetically, controlled
by network activity and by “Darwinian”mechanisms,
was first articulated by Jean-Pierre Changeux and his research
team in 1972.
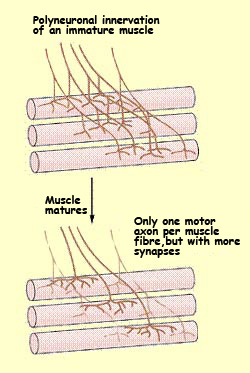
The pioneering studies by Changeux and Danchin revealed
that at the start of development, each muscle fibre is
innervated by axons from several motor
neurons, but by adulthood, it is innervated by the
axon of only one. Similarly, each Purkinje
cell in the cerebellum is innervated by only one climbing
fibre, and the neurons
of the visual cortex, which initially receive connections
from both eyes, eventually retain connections from only
one eye or the other.
Mammals have two types
of muscle fibres, which can be distinguished by their colour
and the speed at which they contract. Fast-contracting
muscle fibres are pale in colour, while slow-contracting
fibres are darker. “Fast-twitch” fibres are
involved in phasic contractions, while “slow-twitch” fibres
are more involved in postural adjustments.
Researchers have also found that the motor neurons that innervate
fast-twitch muscles conduct nerve impulses very rapidly and
can emit action potentials at high frequencies (30 to 60
impulses per second). In contrast, the motor neurons that
innervate slow-twitch muscles conduct impulses more slowly
and can generate only 10 to 20 action potentials per second.
There is thus a clear correspondence between the type of
motor neuron and the type of muscle fibre.
It is known, for example, that newborn kittens have only
slow-twitch muscle fibres, and that these differentiate into
fast-twitch and slow-twitch fibres during the first few weeks
of life. This raises the question of whether it is the muscle
fibres that induce the correct type of corresponding motor
neuron, or whether it is instead the neurons that, by innervating
the muscle fibres, somehow tell them what kind of muscle
fibre to become.
To answer this question, experiments were conducted in which
fast motor neurons were surgically reconnected to slow muscle
fibres, and vice versa. These experiments showed that the
latter hypothesis is correct: it is the nervous system that
controls the contraction speed of the muscle fibres that
it innervates. This implies that the motor neurons are capable
of influencing the expression of the muscle fibres’ genes,
because the contraction speed of a muscle fibre depends on
different versions of myosin light chain
fibres. Part of the reason for this remarkable phenomenon
might be that the differing activity patterns of the two
types of motor neurons have distinct differentiating effects
on the muscle fibres.