The solitary tract nucleus,
in the medulla, also plays a role in the process of falling asleep, through its
projections to the preoptic
area of the hypothalamus. For example, stimulation of the pneumogastric nerves
is known to cause subjects to fall asleep. Also, martial arts practitioners know
how their alertness can be affected by blows to the neck. Likewise, Balinese massage
practitioners are familiar with the relaxing effects of massaging the carotid
area. The word “carotid” itself comes from a Greek word that means
“causing deep sleep”.
Given the great complexity of the
circuits involved in sleep, we can see that the various forms of insomnia can have many
different causes: persistent stimuli that maintain wakefulness, or underfunctioning
of the antiwaking system, or a phase lag in the body’s biological clock.
THE NEURONAL SWITCHES FOR WAKING AND SLEEPING
Once the brain’s
wakefulness
network has been activated, its activity is maintained by internal and external
stimuli. How then does the desire to sleep come about?
First,
of course, these stimuli must subside. But after that, the process of falling
asleep is actually generated by the wakefulness network itself!
These GABAergic neurons, sometimes described as
non-REM (or NREM) sleep-on neurons, are at their most active during deep (non-REM)
sleep and are inactive during wakefulness and REM sleep. Electrical stimulation
of these neurons quickly causes sleep, and their destruction causes insomnia.
This insomnia can be interrupted, however, by the injection of muscimol (a GABA
analogue) into the posterior hypothalamus, where several components of the wakefulness
system converge.
The preoptic area of the hypothalamus
is an ideal location for this sleep-inducing system, because this area is a strategic
crossroads that controls vital functions such as thermoregulation, hunger, and
reproduction. This area can therefore monitor and analyze the body’s functional
status and trigger sleep before the body becomes too fatigued, at the ideal time
indicated by its biological clock.
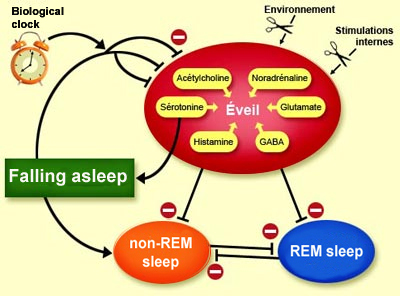
The suprachiasmatic nucleus, which is
the main component of this biological clock, is thus also involved in triggering
sleep. When its neurons are damaged, the normally long periods of wakefulness
shorten and become randomly distributed across the day. These neurons influence
wakefulness through one of their neuropeptides: vasopressin.
(Note that the effects that the vasopressin synthesized by the suprachiasmatic
nucleus has on the brain are completely different from those of the vasopressin
produced by the posterior pituitary gland, which acts mainly on kidney function
and blood pressure.)
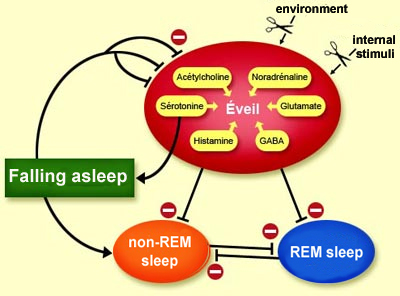
Back to serotonin, however. This neurotransmitter
plays a specific dual role. On the one hand, it is produced in large amounts during
wakefulness and contributes importantly to this state. But on the other hand,
serotonin also plays a fundamental role in the process of falling from wakefulness
into non-REM sleep.The explanation for this contradiction took scientists a while
to find. They had even long regarded serotonin as the “sleep hormone”,
because in animal experiments, destroying the neurons that synthesized it, or
inhibiting its synthesis in other ways, caused periods of sleeplessness that lasted
several days, but the same animals could sleep again if the immediate precursor
of serotonin was then injected into the preoptic area of their anterior hypothalamus.
Since,
then, scientists have obtained a far better understanding of why a lack of serotonin
in the anterior hypothalamus prevents the onset of sleep. This improved understanding
led to the hypothesis that the preoptic area of the anterior hypothalamus did
not operate as a “sleep centre”, but rather as an area that imposed
an inhibition on wakefulness. This hypothesis was subsequently confirmed electrophysiologically.
It was found that the measured unit activity of the serotonergic raphe neurons
is at its greatest during wakefulness, declines at the onset of non-REM sleep,
and ceases during REM sleep. This gradual onset of electrical silence as someone
moves from non-REM sleep into REM sleep thus indicates that the raphe neurons
stop releasing serotonin into the synapses because it has done its work of inhibiting
wakefulness, and its levels can therefore be allowed to decline.
Once
the wakefulness network has been thus inhibited by the antiwaking system, the
pacemaker
cells for non-REM sleep can be expressed. At the same time, the disinhibition
of the thalamic pacemaker also contributes to the onset of sleep. The rhythmic
activity that then becomes established in the thalamus prevents the cortex from
performing cognitive processes that require rapid communication between the thalamus
and the cortex, such as that which takes place during waking and dreaming.
Each of the brain’s different states
of alertness (wakefulness, non-REM sleep, and REM sleep) has its
own distinct oscillation pattern, or rhythm. This rhythm is the result of
interactions between the thalamus and the cortex, which in turn depend on modulations
in the brainstem and the hypothalamus.
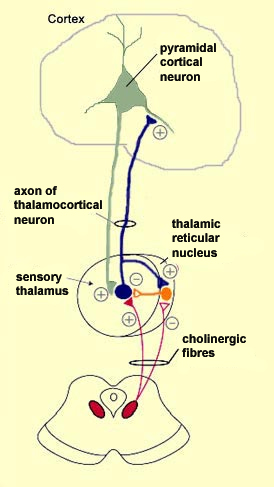
For
example, the neurons of the mesopontine
cholinergic nuclei of the ascending pathway, which are located in the rostral
part of the pons, project their axons to the thalamus. There they make cholinergic
connections not only in its sensory areas, but also in its reticular nucleus,
a layer of neurons that surrounds the thalamus like a skin and exerts a general
inhibiting effect on it by means of the neurotransmitter GABA. (By the way, despite
its name, the thalamic reticular nucleus has nothing to do with the reticular
formation.)
The cholinergic neurons from the pons sensitize the sensory
thalamus but inhibit the reticular nucleus. How can this be so? The answer is
that these two structures have different kinds of receptors for acetylcholine
and hence respond to it differently. The sensory thalamus is sensitized by the
activation of its nicotinic receptors for acetylcholine, whereas
the reticular thalamus is inhibited by the activation of its muscarinic
receptors for this same neurotransmitter.
When the brain
is awake, its cholinergic, histaminergic, and noradrenergic networks thus activate
the thalamus in two ways: directly, by facilitating the sensory thalamus, and
indirectly, by inhibiting the reticular nucleus and thus suppressing its general
inhibiting effects on the thalamus.
It
is important to note that acetylcholine does not excite the neurons of the sensory
thalamus directly. Instead, it simply sensitizes them by depolarizing them slightly,
so that instead of firing action
potentials in bursts, they begin doing so at regular intervals. In this state,
the thalamic neurons are more sensitive to sensory inputs. As a result, the activity
of the pyramidal cortical neurons, which receive major connections from these
thalamocortical neurons, is desynchronized, and the EEG trace becomes typical
of the waking state: low amplitude but high frequency. Note that these pyramidal
cells receive direct nicotinic cholinergic excitation directly from the basal
nucleus of Meynert as well as from the thalamocortical neurons.
During
the minutes when an individual is falling asleep (Stage 1 non-REM sleep), the
firing frequency of the noradrenergic, cholinergic, and serotonergic neurons of
the activating system in the brainstem decreases, so that the thalamus is less
activated.
Concurrently, the inhibition of the neurons
of the thalamic reticular nucleus is correspondingly released, so that they can
resume their spontaneous oscillating activity, which makes their inhibiting effect
on the thalamocortical neurons grow stronger and stronger. The rhythmic action
potentials that are then generated by the GABAergic pacemaker neurons of the reticular
nucleus cause
a cyclical hyperpolarization of the thalamocortical neurons, thus helping
to generate the rhythmic activity of the thalamus. The thalamus then becomes less
and less sensitive to stimuli from the environment, which is the distinctive feature
of the deepest phase of sleep.
In Stage 2 of non-REM sleep, the cortex
goes into an automatic activity pattern of thalamic origin, characterized by sleep
spindles on the EEG. These spindles are caused by the process just described:
the rhythmic firing of the reticular neurons produces cyclical hyperpolarizations
in the thalamocortical neurons, followed by bursts of action potentials. These
potentials are received by the cortical cells, where they generate the sleep spindles.
The slow, high-amplitude waves produced in stages 3 and 4 of non-REM
sleep result from the hyperpolarization of the pyramidal cells of the neocortex,
which is triggered by local GABAergic interneurons, most likely under the influence
of the preoptic neurons of the anterior
hypothalamus. The thalamic neurons, whose membrane
potential is then even more negative than during sleep spindles (seen mainly
in Stage 2) probably also contribute to these slow cortical waves.
Lastly,
during REM sleep, one cause of the desynchronized EEG traces characteristic of
this stage is the influence of the cholinergic neurons on the thalamic cells,
which prevents the expression of their rhythmic oscillatory activity by the same
mechanisms described above with regard to wakefulness.
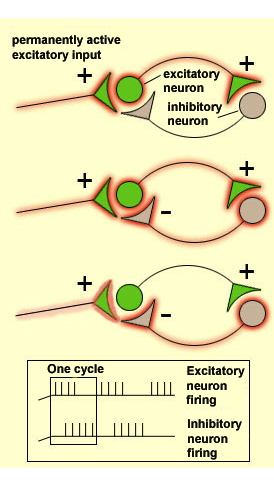
There
are various types of neuronal connectivity that encourage rhythmic bursts of action
potentials within neural networks. One of the simplest is a reciprocal connection
between an excitatory neuron and an inhibitory neuron, both of which are activated
by a third neuron whose own activation pattern may be steady, with no rhythmic
bursts. As long as this continuous activation of the excitatory neuron by the
third neuron persists, the activity of this excitatory neuron will be interrupted
regularly, because this neuron activates the inhibitory neuron, which inhibits
it in return. But then the temporary cessation of the excitatory neuron’s
activity immediately causes the inhibitory neuron’s activity to cease. Hence
the excitatory neuron is once again receptive to the continuous input from the
third neuron. The excitatory neuron is therefore activated again, which quickly
activates the inhibitory neuron, and the oscillatory cycle continues.