|
|
| Funding for this site is provided by readers like you. | |
|
|
|
|
|
|||||
|
|
|||||||
|
|
|
|
| |
Embodied
Cognition and Emotions
| |
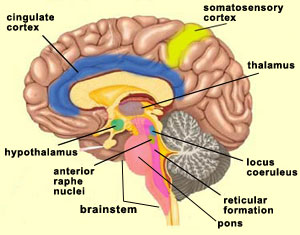
Some neurobiological models of consciousness, such as the global workspace theory, assume that the contents of consciousness are widely distributed in the brain. This assumption has been confirmed by many brain-imaging experiments, in particular those of Stanislas Dehaene and his collaborators. In these experiments, when the amount of time that a word was projected onto a screen was extended just past the threshold required for subjects to perceive it consciously, there was a major increase in activity in their frontal, prefrontal, anterior cingulate, and parietal cortexes. Thus conscious sensory inputs would appear to produce far more extensive brain activity than comparable unconscious stimuli, and a sudden activation of the frontal and parietal lobes would appear to be the typical signature of a conscious perception. But this perceptual consciousness, or as some would call it, primary consciousness, is not the only form of consciousness. When we are trying to associate consciousness with particular structures in the brain, we must therefore clearly define what level of consciousness we are talking about. For example, the first condition necessary for the brain to be able to process external sensory stimuli consciously is that it must be in an appropriate state of alertness (for instance, awake rather than asleep).
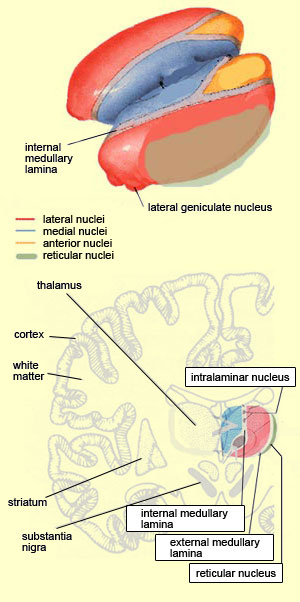
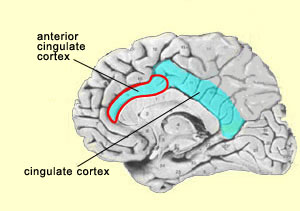
It should be noted here that the activity of the reticular formation, like that of the primary sensory areas, seems to be necessary but not sufficient for a more elaborate level of consciousness. This latter level is attained with what several authors call primary consciousness, meaning a waking state in which we are in relationship with our environment “here and now”. On the basis of the research done by Swedish neuroscientist Bjorn Merker, it seems that the brainstem plays a more important role in primary consciousness than was formerly believed. Damasio calls this type of consciousness “core consciousness” and says that it depends chiefly on the cingulate cortex and on the intralaminar nuclei of the thalamus. Indeed, experiments have shown that bilateral destruction of the centromedial portion of the intralaminar nuclei of the thalamus also eliminates consciousness, produces a coma, or causes other states similar to brain death. In addition, this region of the thalamus is one of the main sites acted upon by anaesthetics and by antipsychotic drugs. Models of consciousness that attribute a role to the thalamus are no recent development. As far back as 1984, Francis Crick offered one of the first hypotheses about consciousness, the “thalamic searchlight hypothesis”, according to which the thalamus controlled which region of the cortex became the focal point for consciousness. A similar but more sophisticated idea has recently been proposed by Rodolfo Llinas. He hypothesizes that the oscillations of certain neurons in the thalamus serve as a sort of basic rhythm with which the cortical oscillations of the various sensory modalities synchronize themselves to form a unified image of the environment—somewhat like an orchestra conductor who provides the beat for all the musicians to follow (see sidebar). This is an original solution to the binding problem.
These thalamocortical loops have come to play an important role in practically all of the neurobiological theories that attempt to explain the higher states of consciousness, for which the lower levels of consciousness that we have been discussing up to now are in a sense only the prerequisites. These higher levels of human consciousness are known as reflexive consciousness and self-consciousness. Reflexive consciousness—this sense that “it is I who am perceiving”—is often presented as a necessary condition for self-consciousness: the feeling of being oneself and not someone else. This autobiographic dimension of consciousness implies that we can form mental representations of conscious experiences in the past or the future, and therefore requires the support of memory and the higher functions that make abstract conceptualization and planning possible. You would therefore expect that the areas of the brain that are known to be involved in these functions, especially in the frontal and parietal lobes, would be actively engaged in self-consciousness. And that indeed has been found to be so in certain studies that addressed this specific question. These higher levels of consciousness also appear to involve other brain structures whose roles were long poorly understood, partly because some of them are located deep in the brain, which made them hard to access. Modern brain imaging techniques have now overcome this problem.
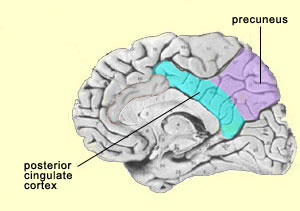
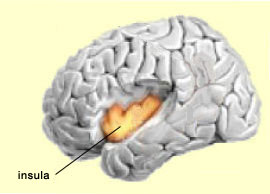
This hypothesis is also consistent with studies that have shown decreased activity in the postero-medial parietal cortex in many states of altered consciousness, such as sleep, anaesthesia, or a vegetative state. Other studies have also shown decreased activity in the precuneus and the posterior cingulate cortex when the subject is under hypnosis, another altered state of consciousness. Lastly, the precuneus also seems to play a role in visual/spatial imagery. For example, some experiments have shown that the precuneus is more active when subjects are is engaged in motor imagery of a finger movement than when they are actually performing this movement. This again seems to indicate that people have a propensity to represent their own bodies in space. The insula (also known as the insular cortex) is another region of the brain that remained little understood for a long time because of its position deep in the folds of the cortex. Also, because it was not associated with the “higher” brain functions, it was of less interest to scientists who were investigating consciousness. But this indifference gave way to intense interest after Antonio Damasio conducted research on the insula and proposed that most of this structure consists of somatic markers. Damasio hypothesized that this part of the cortex maps the bodily states associated with our emotional experiences, thus giving rise to conscious feelings. This hypothesis falls within the school of thought known as embodied cognition, according to which conscious rational thought cannot be separated from emotions and their incarnation in the rest of the body.
The insula has also been convincingly shown to be associated with pain processes, as well as with several basic emotions such as anger, fear, disgust, joy, and sadness. Its most anterior portion is regarded as part of the limbic system. The insula also appears to be deeply involved in conscious desires, such as the active search for food or drugs. What is common to all of these states is that they affect the entire body profoundly—which tends to strengthen the case for the insula’s likely role in the way we represent our bodies to ourselves and in the subjective aspect of emotional experience. Lastly, the insula in humans, and to a lesser extent in the great apes, appears to incorporate two evolutionary innovations that provide these species with a greater ability to read the state of their own bodies than any other mammals. First, the anterior portion of the insula, and more particularly that of the insula in the right hemisphere, is more developed in humans and great apes than in other animal species. This greater development might enable more precise decoding of bodily states—the capability that translates a bad odour, for example, into a feeling of disgust, or the touch of a lover into a feeling of delight. The other major evolutionary modification in the insula is a type of neuron that is found only in the great apes and in humans. These large, elongated, cigar-shaped nerve cells are known as von Economo neurons (VENs). VENs occur only in the insula and in the anterior cingulate cortex. These neurons connect to various parts of the brain, which would be an essential attribute for the higher functions attributed to these two brain structures. When experimental subjects are pricked with a needle, the activity in their cingulate cortex increases; this response is so clear-cut that the neurons in question are often called the “pain neurons”. A fascinating side-note: in 1999, William Hutchison and his colleagues at the University of Toronto showed that these same neurons in the cingulate cortex also become active when the subject sees someone else being pricked with a needle. Thus, for these kinds of neurons, known as mirror neurons, there is no boundary between the self and the other. Primates, including humans, are highly social creatures. Knowing other individuals’ intentions has always been crucial for our survival. That is why we are past masters of the art of internally simulating other people’s minds, perhaps with the help of such mirror neurons. Some neuroscientists, such as V.S. Ramachandran, even suggest that this ability to decode other individuals’ states of mind may even have evolved first, and subsequently been applied to the self, to become what we call self-consciousness. And in Ramachandran’s view, not only the mirror neurons, but all parts of the brain that contribute to language, such as Wernicke’s area in the temporal lobe, must inevitably play a role in this process. This
important role is ascribed to language in several models of higher consciousness,
including that proposed by Michael Gazzaniga, who is known for
his work with “split-brain”patients. But while Gazzaniga's model identifies
the language hemisphere as the locus of this “interpreter” that makes
us conscious of ourselves, other authors, such as Edelman,
argue that consciousness
cannot be attributed to any specific structure in the brain. |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||

| |
|
|
|
|
|
|
|
|