The reticular
formation consists of a disorganized network of
neural fibres with the neurons’ cell bodies scattered
inside of it. Through collateral connections, this structure
receives information from all of the sensory modalities.
When all of these different kinds of information converge
on a single neuron in the reticular formation, they lose
the specificity of their origin and acquire the non-specific
property of activating the excitatory neurons of the wakefulness
network.
Other reticular cells increase muscle tonus and perform the
work that is necessary but not sufficient for attention, learning,
and voluntary
movements.
The reticular formation can thus be
regarded as a sort of filter: out of all the various visual,
auditory, and somatosensory stimuli that surround us all
the time, it allows to pass through to consciousness only
the infinitesimal fraction that are the most immediately
useful to us, or the most intense.
For example, if the noise from your
computer fan is persistent but not too loud, after a while
you simply stop noticing it. You can thus see why, without
the reticular formation, it would be pretty hard to fall
asleep in places that were noisy, uncomfortable, or brightly
lit.
THE BRAIN STRUCTURES
THAT WAKE YOU UP AND PUT YOU TO SLEEP
The cortical activation necessary for you
to be awake is made possible by an “executive
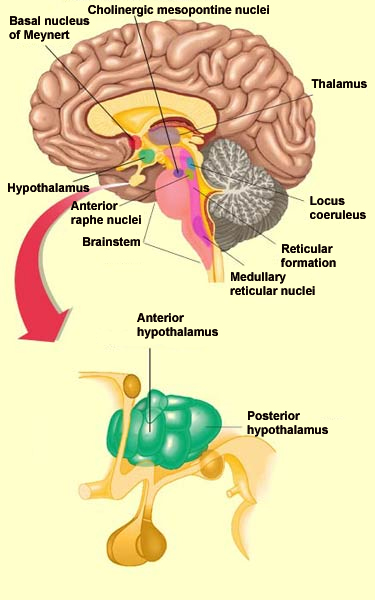
network” for wakefulness, consisting of the posterior
hypothalamus, the intralaminar thalamus, and the basal telencephalon.
But this network is itself influenced by a fairly complex
network of about 10 structures ascending from the lower
brainstem to the basal telencephalon and thereby controlling wakefulness.
Very schematically, the components of these ascending
modulatory systems can be divided into two major pathways,
one ventral, the other dorsal, both of which arise from a part
of the reticular nucleus in the medulla oblongata.
The ventral pathway is called
the reticulo-hypothalamic-cortical pathway. It
projects to the posterior hypothalamus and to the nucleus of Meynert,
which consists of cholinergic neurons and is located in the basal
telencephalon.
The dorsalpathway is
called the reticulo-thalamic-cortical pathway. It
activates the cholinergic mesopontine nuclei, the aspartergic and
glutamergic neurons of the midbrain reticular formation, and the
thalamus.
In addition to maintaining wakefulness, several
of the nuclei of these two pathways use acetylcholine and glutamate
as neurotransmitters and are partly responsible for the cortical
activation that occurs during REM
sleep.
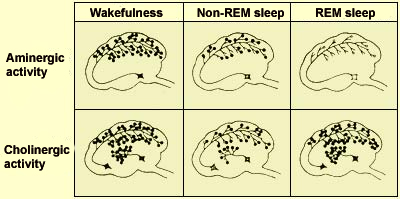
The numbers of
black dots are proportional to the activity of the aminergic
and cholinergic systems during the various stages of wakefulness
and sleep.
Here are descriptions of some of the most
important brainstem nuclei involved in the wakefulness system.
The midbrain
reticular formation projects massively into the
thalamic nuclei, which in turn influence the entire cortex.
The role of this formation is to desynchronize the cortex
in the broad sense, thus facilitating not only wakefulness
but REM sleep as well. Formerly known as the ascending
activating reticular system, it is now regarded simply
as part of the wakefulness network.
The cholinergic mesopontine nuclei also
project to the thalamus. The acetylcholine
produced by these nuclei has two effects: it reduces
the activity of the thalamic reticular nucleus, which is
part of the sleep system, and it activates the thalamocortical
neurons involved in wakefulness.
The magnocellular medullary reticular nuclei,
whose neurons are cholinergic or aspartergic/glutamergic,
are the origin of both the reticulo-thalamic-cortical pathway
and the reticulo-hypothalamic-cortical pathway. Their projections
therefore run to the midbrain reticular formation and the
cholinergic mesopontine nuclei, as well as to the basal telencephalon
and the posterior hypothalamus.
The nuclei of the locus
coeruleus are located in the dorsal part of the
pons, and their noradrenergic projections influence brain
structures such as the thalamus, the hippocampus, and the
cortex. The locus coeruleus is at its most active when an
individual is awake and active. It is less active during
calm wakefulness, even less active during non-REM sleep,
and completely quiescent during REM sleep.
The serotonergic nuclei of the
anterior raphe (also known as the superior raphe)
send serotonin to the hypothalamus and the cortex. These nuclei
are active during wakefulness. Their overall effect is to support
wakefulness, and, unlike with other groups of aminergic neurons,
lesions to these nuclei not only do not cause even transient
sleepiness, but actually cause prolonged insomnia that lasts
several days. The likely reason for this apparent contradiction
is that this system, which innervates the anterior hypothalamus
both in the preoptic area and in the circadian clock circuits
of the suprachiasmatic nucleus, seems to measure the duration
and intensity of wakefulness. Wakefulness might thus eventually
cause its own inhibition through a negative feedback loop.
In other words, having
been awake for a certain time leads to sleep.
All of these structures in the brainstem
receive collateral projections from the sensory and vegetative
inputs which thus help to maintain their activity (see sidebar).
Thus we are talking about a complex network in which the pharmacological
excitation of one component leads to the activation of all the
others. This redundant arrangement also explains why the deactivation
of a single system is followed, after a few days, by a complete
recovery of wakefulness. Thus, taken in isolation, none of the
structures just described is indispensable for the activation of
the cortex.
Researchers can generate
PGO waves during wakefulness, rather than during REM sleep,
by exposing their subjects to sudden, strong stimuli that
startle them. This suggests that the PGO waves that arise
spontaneously during REM sleep may be generated by the internal
activation of the neuronal circuitry of the startle reflex.
The activity of the
pontine inhibitory neurons affects not only the motor neurons,
but also the nuclei of the dorsal columns, where, during
REM sleep, these pontine neurons reduce responsiveness to
somaesthetic stimuli.
One of these characteristics consists of the rapid
eye movements that occur during REM sleep and from
which it gets its name. Their purpose is unknown, but it is known
that the signals generating them originate in the pontine
reticular formation and are transmitted to the motor
layers of the superior
colliculi. The collicular neurons in turn send projections
to the paramedian pontine reticular formation (PPRF), which co-ordinates
the duration and direction of these eye movements.
Another singular characteristic of REM sleep whose source has
been located is the near-total
bodily paralysis that accompanies it. The intense neural
activity observed during REM sleep excites the vast majority
of the neurons in the cortex, including those in the primary
motor cortex. These motor neurons thus generate organized
sequences of activities that represent commands for bodily movements.
But during REM sleep, only the respiratory muscles and the muscles
of the eye and the middle ear will actually be able to carry
out these commands–they never reach the motor neurons of
the arms and legs.
During REM sleep, the increased activity of cholinergic
neurons in the pons excites other, glutamergic neurons
in the pontine reticular formation. These neurons
in turn send projections to and activate interneurons in the
magnocellular reticular nuclei of the medulla. The axons of these
interneurons descend into the spinal cord, where they release
glycine, thus strongly inhibiting the motor neurons by hyperpolarizing
them.
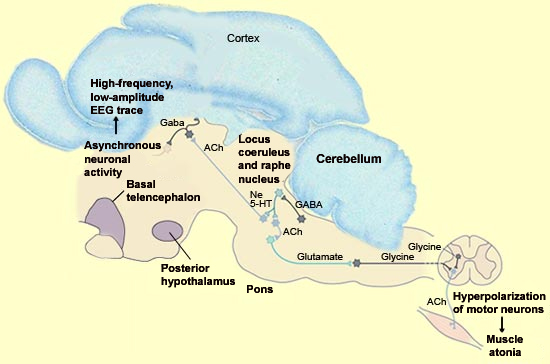
Sagittal section
of the brain of a cat, the model of choice for studying REM sleep,
with a simplified diagram of certain structures involved in controlling
REM sleep(Ne = norepinephrine, 5-HT
= serotonin, ACh = acetylcholine)
Source: Adapted from Principles
of Neural Science, Kandel,
Schwartz, and Jessell, Eds.,
Appleton & Lange, Pub.
Another feature of REM sleep, the pontogeniculooccipital
(PGO) spikes that occur intermittently during it,
also originate in the pontine reticular formation.
They then propagate via the thalamus to the occipital
cortex, though as their name indicates, they are more
readily detectable in the lateral
geniculate nuclei, which act as relays in the brain’s
visual system.
PGO waves are among the various phasic
events that occur during REM sleep, along with the rapid eye
movements and changes in breathing and heart rates. PGO waves
can be generated in the absence of REM sleep by stimulation of
the pons with acetylcholine, especially in the peribrachial area
of the pons. It is in this reticular area, around the superior
cerebellar peduncle and underneath the locus coeruleus, that
PGO waves are generated.
Many of these neurons that project to the thalamus are cholinergic.
They briefly fire action potentials just before each PGO wave
on the ipsilateral side of the brain.
The serotonin in the raphe system inhibits PGO waves by hyperpolarizing
the cells that generate them. We can thus understand why the
halt in the activity of the serotonergic cells during the transition
from non-REM sleep to REM sleep triggers PGO waves.