|
|
| Funding for this site is provided by readers like you. | |
|
|
|
|
|
|||||
|
|
|||||||
|
|
|
|


|
|
| |
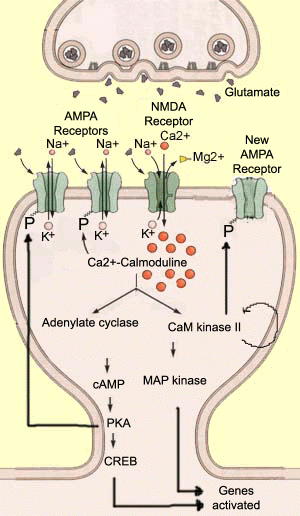
Long-term potentiation (LTP) is a process in which synapses are strengthened. It has been the subject of much research, because of its likely role in several types of memory. LTP is the opposite of long-term depression (LTD). In LTP, after intense stimulation of the presynaptic neuron, the amplitude of the post-synaptic neuron’s response increases. The stimulus applied is generally of short duration (less than 1 second) but high frequency (over 100 Hz). In the postsynaptic neuron, this stimulus causes sufficient depolarization to evacuate the magnesium ions that are blocking the NMDA receptor, thus allowing large numbers of calcium ions to enter the dendrite. These calcium ions are extremely important intracellular messengers that activate many enzymes by altering their conformation. One of these enzymes is calmoduline, which becomes active when four calcium ions bind to it. It then becomes Ca2+/calmodulin, the main second messenger for LTP. Ca2+/calmodulin then in turn activates other enzymes that play key roles in this process, such as adenylate cyclase and Ca2+/calmodulin-dependent protein kinase II (CaM kinase II). These enzymes in turn modify the spatial conformation of other molecules, usually by adding a phosphate ion to them. This common catalytic process is called phosphorylation.
To give some idea of the complexity of the metabolic sequences responsible for LTP, we will mention three of the other enzymes currently being studied. Protein kinase C (PKC) appears to phosphorylate AMPA receptors at the same site as CaM kinase II. Inhibitor 1 (ou I1) seems to be activated by PKA and prevent phosphatase 1 from dephosphorylating AMPA receptors. And tyrosine kinase SRC may be activated directly by the AMPA receptors, and then phosphorylate the NMDA receptors.
In addition to all of the post-synaptic mechanisms involved in the establishment of LTP, it has long been postulated that some presynaptic modifications occur during the ensuing maintenance phase. But certain modifications, such as an increase in the amount of glutamate released by the presynaptic neuron, would imply the presence of a retrograde messenger that goes back to this neuron and modifies it. Because nitric oxide (NO) is a gas in its natural state, and can thus diffuse through cell membranes, it would be an ideal candidate for this role. But its involvement is still the subject of much debate and controversy. |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||

| |
|
|
|
|
|
|
|
|