There are many neurotransmitter
systems that can influence anxiety. In addition to GABA, serotonin is
often cited for its high levels in certain parts of the brain
associated with anxiety. Serotonin is also known for its
role in obsessive-compulsive
disorder and depression,
both of which are closely related to anxiety. And the fact
that serotonin-reuptake-inhibiting
antidepressants have an effect on these conditions shows
that serotonin probably plays a role in them.
Because GABA is the primary inhibitory
neurotransmitter in the brain, it obviously plays an important role
in controlling the neuronal hyperactivity associated with anxiety.
Since GABA agonists can induce comas, the pharmaceutical industry
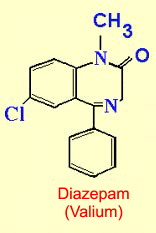
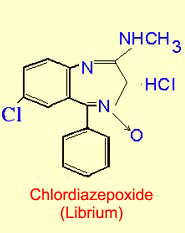
has had to turn to other ligands that enhance GABA’s effects. Benzodiazepines such
as Valium and Librium, which act as modulators for GABAa
receptors, have become some of the best anxiolytics available.
In just the
same way that, once scientists had characterized the receptors
for opiates in the brain, they also discovered natural endogenous
morphines that bind to them, researchers have now identified
molecules produced by the body that bind to exactly the same
site on the GABAa receptor as synthetic benzodiazepines. These
endogenous benzodiazepines, or endozepines, which seem
to be produced chiefly by the glial
cells, have been partially purified in the human brain.
The term “endozepines” designates both diazepam-binding
inhibitor (DBI) and the peptides derived from it, including
triakontatetraneuropeptide (TTN) and octadecaneuroepetide (ODN).
Despite the potential importance of endozepines
as endogenous ligands for the benzodiazepine site on GABAa receptors,
very little research has been done on the role of these peptides.
Endozepines may of course achieve some of their effects by modulating
GABAa receptors, but other effects, such as the anorexigenic effect
of ODN, might involve a separate metabotropic
receptor.
DBI may be an inverse agonist for the benzodiazepine
site on the GABAa receptor. In other words, DBI may reduce
the receptor’s chloride permeability and hence GABA’s
effectiveness, and thus would be anxiogenic. In fact, DBI is
a disconcerting molecule for neurobiologists, who have shown
far less enthusiasm about endozepines than they have about enkephalins
and endorphins. Nevertheless, these endogenous benzodiazepines,
through their modulating effects on GABA, are thought to enable
it to play a more flexible role in neurophysiological processes.
It is also thought that disturbances in their activity may play
a role in chronic anxiety.
The molecules conventionally identified
as neurotransmitters are not the only ones that may have
an effect on anxiety. Neuropeptides such
as cholecystokinin (CCK) may also be anxiogenic; the release
of this molecule may be enhanced by serotonin and norepinephrine
in the cortico-limbic system. The use of cholecystokinin
antagonists as anxiolytics is therefore being considered.
Another peptide, CRH, is a powerful
anxiogenic whose release is stimulated by stress. Neuropeptide
Y has an anxiolytic effect almost as powerful as the benzodiazepines’.
It is believed that under normal conditions, CRH and neuropeptide
Y, through their opposing effects, constitute a system that
controls the integration of stress signals in the amygdala.
GABA RECEPTORS
GABA (gamma-aminobutyric acid) exerts
its effects through at least three different types of receptors:
the GABA-A receptor (which is the best known), and the GABA-B and
GABA-C receptors. The GABA-A and C receptors are ionotropic,
while the GABA-B receptor is a metabotropic
receptor that modulates the opening of potassium channels through
second messengers involving a G-protein.
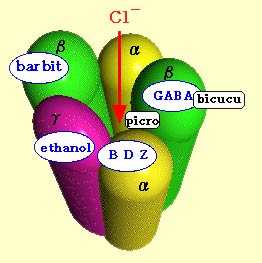
Each
of these receptors is a macromolecular complex comprising several
sub-units. For example, the GABA-A receptor is composed of
5 sub-units surrounding a channel that is preferentially
permeable to chloride ions and to a lesser extent to bromide
ions. The GABA receptor site appears to be located in the large
extracellular domain of the beta sub-unit. These 5 sub-units
have 16 known isoforms, each produced by a different gene.
In addition to the primary binding sites
for GABA, the GABA-A receptor has other secondary binding sites
for molecules that modulate GABA’s effects, such as benzodiazepines,
barbiturates, convulsants, steroids, and alcohol.
These modulating agents alter GABA’s
efficiency by inducing a change in the protein architecture
of the GABA-A complex. This change modifies the size of the
channel, which in turn modifies the receiving neuron’s
permeability to chloride ions. Since chloride ions are negatively
charged, when they enter the neuron, they hyperpolarize it.
The result is an inhibition of neuronal
activity and a general anxiolytic effect.Treatment with benzodiazepines
thus helps to reduce anxiety by potentiating the effect of GABA, and
more specifically, by making the chloride channel open more frequently.
However, in the absence of GABA at the primary site on the GABA-A receptor,
the modulating molecules have no effect on the neuron’s chloride
permeability.