|
|

 |
|

|
|

|
An amazing discovery in the 1970s
demonstrated that a rat’s hippocampus is a veritable
spatial map of the environment through which it moves. Certain
pyramidal neurons in area CA1 of the rat hippocampus become
active only when the rat is located in a specific part of
its environment.
There are 1 million of these “place
cells” in area CA1 of the rat hippocampus, so that
if each one is assigned a specific point in space, together
they can form a very precise cognitive map that tells the
animal where it is at any given time. Moreover, when a rat
explores a new environment, it forms a new cognitive map
that can be very stable, lasting weeks or months.
According to O’Keefe and Nadel,
the researchers who discovered the existence of these cognitive
maps, one of their functions might be to provide a context
to which memories can be attached. An event recorded in memory
could thus be made autobiographical (situated in time and
space). This would explain the fundamental role that the
hippocampus plays in episodic
memory in human beings.
|
|
|
| PLASTICITY IN NEURAL
NETWORKS |
|
No one neuron alone contains all the information
needed to reconstruct a memory. Rather, the trace of that memory
is latent or virtual. Its existence can be manifested only when a
network of many interconnected neurons is activated.
Multiple memories can be encoded within a
single neural network, by different patterns of synaptic connections.
Conversely, a single memory may involve simultaneously activating
several different groups of neurons in different parts of the brain.
This association of groups of cortical neurons
distributed across different parts of the brain is made possible
by certain networks of neurons that are pre-wired for this task.
Certainly the best known of these networks are the circuits of
the hippocampal formation, which are involved in establishing explicit
long-term memories.
The information from the visual, auditory,
and somatic associative cortexes arrives first at the parahippocampal
region of the cortex, then passes through the enthorinal cortex
and on to the hippocampus proper.
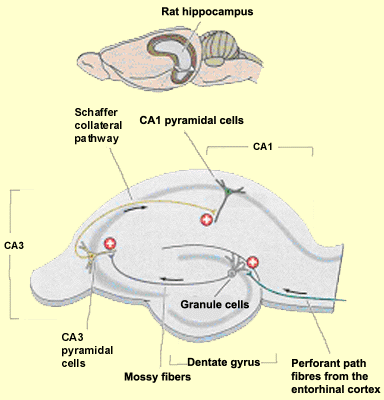
Within the hippocampus, the information passes through three distinct
regions in succession.
The hippocampus proper is composed of regions
with tightly packed pyramidal neurons, mainly areas CA1, CA2, and
CA3. (“CA” stands for Cornu Ammonis, or Horn of Ammon.
The reference is to the ram’s horns of the Egyptian god Ammon,
whose shape these three areas together roughly resemble.) This
is what is called the trisynaptic circuit or trisynaptic loop
of the hippocampus.
| Information
enters this one-way loop via the axons of the entorhinal cortex,
known as perforant fibres (or the perforant path, because it
penetrates through the subiculum and the space that separates
it from the dentate gyrus). These axons make the loop’s first
connection, with the granule cells of the dentate gyrus.
From these cells, the mossy fibres in turn project to make
the loop’s second connection, with the dendrites
of the pyramidal cells in area CA3.
The axons of these cells divide into two branches. One branch
forms the commissural fibres that project to the controlateral
hippocampus via the corpus callosum. The other branch forms
the Schaffer collateral pathways that make the third connection in
the loop, with the cells in area CA1. |
 |
It is in these synapses that the spatial
memory associated with the hippocampus seems to be encoded (see
sidebar). This region also displays a high propensity for long-term
potentiation (LTP), though this same phenomenon is also observed
in many other parts of the hippocampus as well as in the cortex.
Lastly, the axons of the cells in CA1 project
to the neurons of the subiculum and of the entorhinal cortex. The
receiving portion of the hippocampal formation thus consists of
the dentate gyrus, while the sending portion consists of the subiculum.
The axons of the large pyramidal neurons of the subiculum then
project to the subcortical nuclei via the fimbria, a thin tract
of white matter at the inner edge of the hippocampus. Lastly, the
information returns to the sensory cortical areas from which it
came before it was processed by the hippocampus. |
|